the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 14 May 2020
| 14 May 2020
An alternative interpretation of Peltochelys duchastelii as a paracryptodire
Walter G. Joyce
Yann Rollot
Peltochelys duchastelii is an enigmatic turtle from Early Cretaceous (middle Barremian–early Aptian) Sainte-Barbe Formation of Bernissart, Belgium. In more recent literature, there has been strong support for the trionychian affinities of this taxon, but this interpretation is less consistent with external data, in particular biogeography and temporal considerations. We provide a reinterpretation here of the lectotype of Peltochelys duchastelii that differs from more recent ones in key features. Although we agree that this turtle lacks mesoplastra, we conclude that it has 11 pairs of peripherals (not 10), likely had 12 pairs of marginals (not 11), lacks a contact between peripheral 1 and costal 1, and that a midline contact of peripherals 1 hinders the nuchal from contributing to the margin of the shell. This unusual set of characters is otherwise known from roughly coeval taxa from Europe and North America. Phylogenetic analysis yields a previously unrecognized clade of basal paracryptodiran turtles from the Late Jurassic to Paleogene of Euramerica united by a nuchal that is withdrawn from the anterior margin of the shell. The name Compsemydidae is referred to this clade. The novel hypothesis suggests that Peltochelys duchastelii did not immigrate from Asia but instead originated in Europe.
- Article
(5484 KB) - Full-text XML
-
Supplement
(481 KB) - BibTeX
- EndNote




Peltochelys duchastelii Dollo, 1884, is a fossil turtle from the Early Cretaceous of Belgium with a particularly complex history of research. The species was originally named by Dollo (1884) based on three individuals, in particular the shell of an adult and two partial skeletons of juveniles. All specimens were collected in Early Cretaceous (middle Barremian–early Aptian) exposures of the Sainte-Barbe Formation (Yans et al., 2005) in the underground coal mines of Bernissart, located in Hainault Province, Belgium. Dollo (1884) only provided an extremely brief description of his new species that highlights the presence of a single gular scute (his plaque intergulaire), the absence of mesoplastra, and a neck inconsistent with cryptodiran neck retraction.
Lydekker and Boulenger (1887) concluded that the type material of Peltochelys duchastelii is referable to the Early Cretaceous (Valanginian) taxon Tretosternon (their Tretosternum) bakewelli (Mantell, 1833), together with a small assortment of specimens from the Early Cretaceous (Berriasian) of Sussex, United Kingdom, and the Early Cretaceous (Barremian) of the Isle of Wight, United Kingdom. It is difficult to parse out the rationale of these authors (Joyce, 2017), as the available material shows little anatomical or temporal overlap. This synonymy was nevertheless accepted by subsequent authors (e.g., Lydekker, 1889; Dollo, 1909; Wegner, 1911), and Tretosternon bakewelli was typically characterized by reference to Peltochelys duchastelii (e.g., Lydekker and Boulenger, 1887; Lydekker, 1889). Nopcsa (1928a, b) concluded that the abovementioned English material can be united with other material from the Cretaceous of North America and Europe into a previously unrecognized group, Helochelydridae (his Helochelydrinae, also known as Solemydidae), based on the presence of a distinct shell texture consisting of raised tubercles, and that Peltochelys duchastelii can be distinguished from helochelydrids by its subdued shell sculpturing and the absence of mesoplastra. Even though this conclusion was further cemented by the influential work of Hummel (1929, 1932), the synonymy of Tretosternon bakewelli with Peltochelys duchastelii was revived once again many decades later by Młynarski (1976, fig. 60.1), who presented the shell of Peltochelys duchastelii under the name Tretosternon bakewelli.
Dollo (1884) originally based Peltochelys duchastelii on three specimens, of which two are juveniles. A century later, Meylan (1988) designated the adult specimen as the lectotype, as he noted that a previously undescribed third juvenile differs from the adult in the number of peripherals. In particular, he noted that the third juvenile has 11 pairs of peripherals, of which the most anterior closely approximate each other, while the adult appears to have only 10 pairs of peripherals. Pérez-García (2011) agreed with this conclusion but noted that the unfigured juvenile of Meylan (1988) appears to be lost. Pérez-García (2015) soon after described two additional juvenile specimens but concluded that all available juvenile material, to the exclusion of the now lost specimen of Meylan (1988), may reasonably represent Peltochelys duchastelii after all.
Karl et al. (2012) expanded the known stratigraphic range of Peltochelys duchastelii by referring new and previously described specimens from the Late Jurassic (Kimmeridgian–Tithonian) of Guimarota and Lourinha, Portugal, and the Early Cretaceous (Berriasian) of Schaumburg-Lippe and Bückeberg, northern Germany. These referrals, however, were convincingly rebuffed by Pérez-García (2014) and Pérez-García et al. (2015b), who noted that the Portuguese material is only diagnostic of an indeterminate pleurosternid, while the German material represents the pleurosternid Dorsetochelys typocardium. Peltochelys duchastelii is therefore still only known from its type locality.
The phylogenetic relationships of Peltochelys duchastelii have been under debate ever since it was discovered. Dollo (1884) referred Peltochelys duchastelii to Pleurodira (his Chelydes), likely based on the purported presence of a single gular (his plaque intergulaire) and the perceived lack of cryptodiran neck retraction. Lydekker and Boulenger (1887), in contrast, concluded that Tretosternon bakewelli (to the inclusion of Peltochelys duchastelii) has affinities with Chelydridae, but, as outlined above, their character evidence was drawn from what is now understood to be a chimera (Joyce et al., 2014; Joyce, 2017). Nopcsa (1928a, b) referred Peltochelys duchastelii to Dermatemydidae (his Dermatemydinae) but did not discuss character evidence that would support this assessment. Hummel (1929, 1932) noted that Peltochelys duchastelii shows dermatemydid characters (i.e., presence of scutes and two suprapygals) and carettochelyid characters (i.e., overall size and shape of the shell, development of a preneural, and the presence of a textured shell) and therefore suggested that it might be the Early Cretaceous dermatemydid ancestor of carettochelyids. Chkhikvadze (1975) referred this turtle to Adocidae, while Nessov (1977) referred it to Nanhsiungchelyidae. Broin (1977) noted that Peltochelys duchastelii occurs too late in the fossil record to be considered an ancestral carettochelyid. Using cladistic arguments, Meylan (1988) hypothesized that Peltochelys duchastelii is the immediate sister to Trionychia based on the presence of 10 pairs of peripherals and paired ventral processes on the nuchal, two characteristics that are otherwise only known to occur in combination in carettochelyids. All subsequent analyses agree that Peltochelys duchastelii is a pan-trionychian, but there is minor disagreement as to whether it is the most basal representative of the lineage (e.g., Brinkman, 1998), the immediate sister to Trionychia (Meylan and Gaffney, 1989; Brinkman and Peng, 1996; Joyce, 2007; Perez-García, 2011), or placed within Carettochelyidae (e.g., Lapparent de Broin, 2001). As the basal evolution of Pan-Trionychia otherwise mostly occurred in Asia, either placement implies independent dispersal of the Peltochelys duchastelii lineage from Asia to Europe (Danilov, 2005). Finally, Danilov et al. (2017) summarized this uncertainty by classifying Peltochelys duchastelii as a eucryptodire of uncertain affinities.
Peltochelys duchastelii is arguably the best figured Early Cretaceous turtle worldwide. Dollo (1884, pl. 2) furnished photographs of the full type series in dorsal and ventral view. Hummel (1929, fig. 2) provided interpretive line drawings of the lectotype in dorsal and ventral view, which were redrawn by Młynarski (1976, fig. 60.1), though under the name Tretosternon bakewelli. Meylan (1988, figs. 3, 4) created new photographs and interpretative line drawings of the lectotype, of which the line drawings were redrawn by Danilov (2005) and Karl et al. (2007). The full type series was once again rephotographed and re-illustrated by Pérez-García (2011, figs. 1–3). Although all available illustrations of the lectotype greatly resemble one another, they differ in regard to some key features of great systematic value. The purpose of this contribution is to provide yet another interpretation of the morphology of Peltochelys duchastelii and to propose the novel hypothesis that this enigmatic turtle is an unusual paracryptodire, not a representative of the pan-trionychian lineage.
-
Testudinata Batsch, 1788
-
Paracryptodira Gaffney, 1975
-
Compsemydidae Pérez-García et al., 2015b
-
Peltochelys duchastelii Dollo, 1884
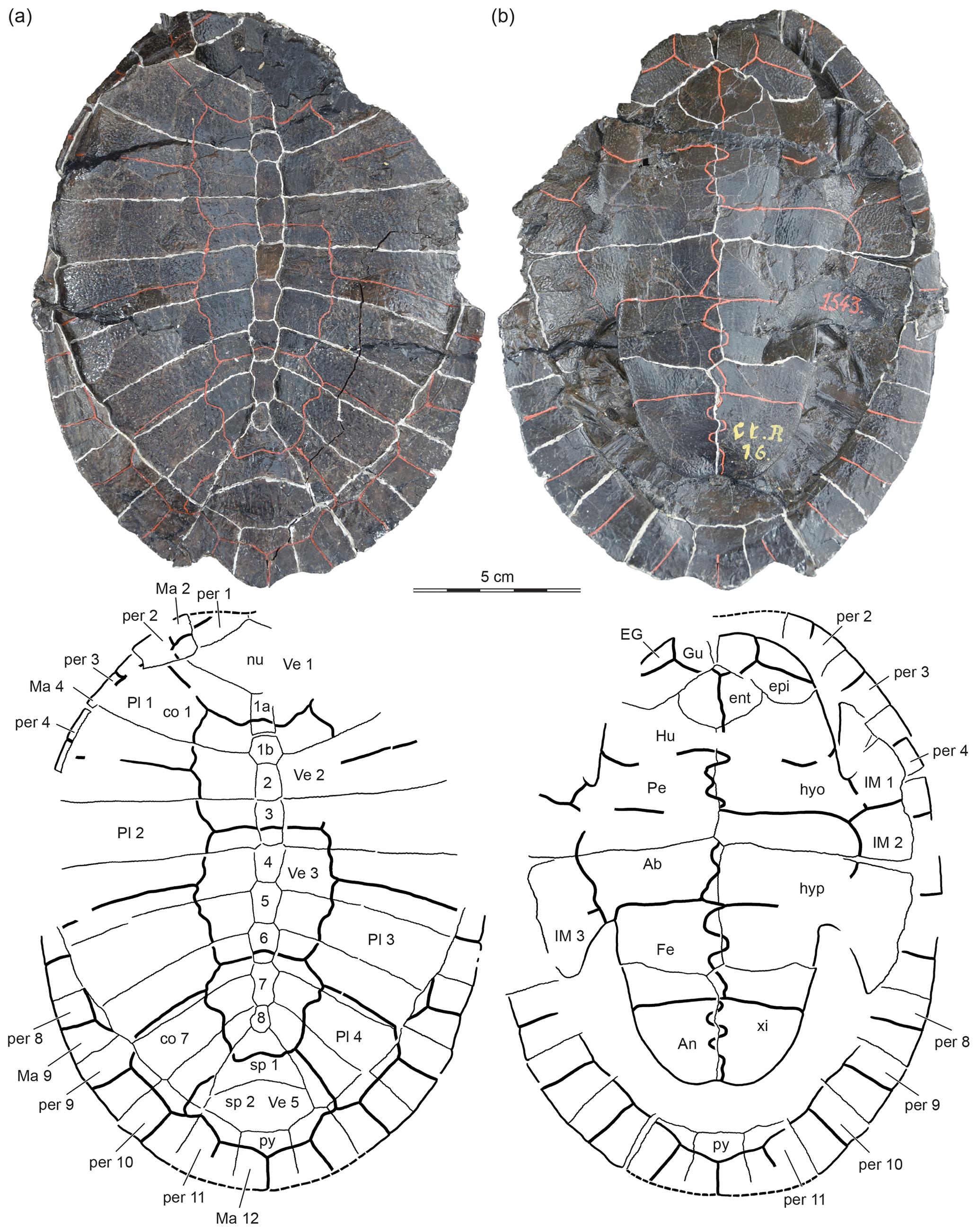
Figure 1IRSNB Ct.R.16, lectotype, Peltochelys duchastelii, Early Cretaceous (middle Barremian–early Aptian) Sainte-Barbe Formation of Bernissart, Hainault Province, Belgium. Photographs and line drawings in (a) dorsal and (b) ventral view. Arabic numerals denote the neural elements. Abbreviations: Ab, abdominal scute; An, anal scute; co, costal; EG, extragular; ent, entoplastron; epi, epiplastron; Fe, femoral scute; Gu, gular scute; Hu, humeral scute; hyo, hyoplastron; hyp, hypoplastron; IM, inframarginal scute; Ma, marginal scutes; nu, nuchal; Pe, pectoral scutes; per, peripheral; Pl, pleural scutes; py, pygal; sp, suprapygal; Ve, vertebral scute; xi, xiphiplastron.
Type specimen
IRSNB Ct.R.16, lectotype, a near complete shell (Dollo, 1884, pl. 2.1–2; Hummel, 1929, fig. 2; Meylan, 1988, figs. 3, 4; Pérez-García, 2011, fig. 1; Fig. 1).
Type locality and stratum
Bernissart, Hainault Province, Belgium (Dollo, 1884); Sainte-Barbe Formation, Early Cretaceous (middle Barremian–early Aptian) (Yans et al., 2005).
Diagnosis
Peltochelys duchastelii can be diagnosed as a representative of Compsemydidae, a clade of paracryptodires, by the presence of a finely textured shell, a sutured bridge, and a sinuous plastral midline sulcus; absence of a contact between peripheral 1 and costal 1; and the withdrawal of the nuchal from the anterior margin of the shell resulting from a midline contact of peripherals 1. Within Compsemydidae, Peltochelys duchastelii is most readily diagnosed by a clear contact of costal 8 with peripheral 11 (likely absent in Riodevemys inumbragigas), a point contact of vertebral 1 with marginal 3 (absent in all others), the absence of mesoplastra (present in all others), a shallow xiphiplastral notch (deep in Toremys cassiopeia and Compsemys spp.), and the presence of a single gular (likely double in all others).
Referred material
Four juvenile specimens, IRSNB Ct.R.17, 18, 291, and 292 (Dollo, 1884, pl. 2.3–6; Pérez-García, 2011, figs. 2, 3; Pérez-García, 2015, figs. 1, 2) from the type locality.
Description
Although the lectotype of Peltochelys duchastelii has already been photographed and/or illustrated independently four times (Dollo, 1884; Hummel, 1929; Meylan, 1988; Pérez-Garcia, 2011) and succinctly described three times (Hummel, 1929; Meylan, 1988; Pérez-Garcia, 2011), we provide yet another visual documentation of this specimen (Fig. 1) but focus on highlighting differences in interpretation.
Carapacial bones
The lectotype of Peltochelys duchastelii possesses nine neural elements (Fig. 1). Hummel (1929) noted that a crack runs through the anterior part of the shell and that the two anterior elements may represent a single damaged neural. The alternative interpretation is the presence of a preneural or the presence of a well-formed supernumerary neural. We are unable to weigh in favor of either hypothesis because all purported sutures are covered in paint, obscuring the true nature of the margins of these elements. To maintain the homology of the more posterior elements, we designate the two anterior ones as neural 1a and 1b (Fig. 1). Neural 7 displays an unusual shape, which is suggestive of two fused neural elements but also consistent with a partially obscured regular neural. We therefore follow previous authors by counting this as a single element. Our final count of eight neurals and a potential preneural matches that of Hummel (1929) and Meylan (1988) but differs from that of Pérez-García (2011), who counted nine neurals, but these differences are semantic and not based on a deeper sense of homology.
The preneural and neural 1 jointly form a hexagonal element with short posterior sides. The shape of neurals 2 and 3 is somewhat unclear, as the original contacts are obscured by movement, but we believe it is most likely that neural 2 is rectangular, while neural 3 is hexagonal with short anterior sides. Neurals 4–6 are clearly hexagonal with short anterior sides, neural 7 is octagonal with short anterior and posterior sides, and neural 8 is an irregular element with four contacts with the surrounding bones.
There is agreement that the lectotype of Peltochelys duchastelii includes eight pairs of costal elements that lack a midline contact (Fig. 1). As a trend, these elements decrease in anteroposterior length from costal 1 to costal 8. The anterior two elements expand distally and are oriented to the anterior. Costal 3 is rectangular and oriented laterally. The posterior five costal elements are slightly expanded distally and are increasingly oriented posteriorly. In our interpretation of the peripheral series (see below), costal 1 lacks a contact with peripheral 1 but contacts peripherals 2 and 3. The remaining costals each have two contacts with the remaining peripheral series, including a clear contact of costal 8 with peripheral 11, as documented on the right side of the specimen.
The lectotype of Peltochelys duchastelii possesses two distinct suprapygal elements (Fig. 1). Suprapygal 1 is a large, trapezoidal element with a short anterior contact with neural 8, two elongate lateral contacts with costal 8 and a broad, convex posterior contact with suprapygal 2. Suprapygal 2 is a lenticular element that has a convex anterior contact with suprapygal 1, a short anterolateral contact with costal 8, and three similarly sized posterior contacts with peripherals 11 and the pygal. The pygal is rectangular.
Substantial differences exist in the interpretation of the nuchal and the peripheral series, which we justify more extensively in the Discussion below. We side with Dollo (1884) and Hummel (1929) by recognizing the presence of 11 pairs of peripherals, but we go further by concluding that the nuchal did not contribute to the margin of the shell. In our interpretation, the nuchal is a heptagonal element that has a broad anterolateral contact with peripheral 1, a short lateral contact with peripheral 2, an elongate posterolateral contact with costal I, and a short posterior contact with neural 1a. The point at the anterior margin of the slightly displaced nuchal suggests that this element did not contribute to the anterior margin of the shell. The alternative interpretation of Meylan (1988) and Pérez-García (2011) suggests that the nuchal contributes broadly to the anterior margin of the shell, the plesiomorphic condition for turtles. In contrast to Meylan (1988), we are unable to find paired nuchal processes in the lectotype of Peltochelys duchastelii, in part because the visceral side of the nuchal is covered from view. This agrees with the observations made by Pérez-García (2011), who also was unable to find any structures on the visceral side of the nuchal.
In our interpretation of the peripheral series, peripheral 1 is damaged, but the remaining triangular part of the bone highlights a broad posteromedial contact with the nuchal, the absence of a contact with costal 1, and a clear posterolateral contact with peripheral 2 (Fig. 1). The pointed tip of the nuchal suggests that peripheral 1 had a midline contact with its counterpart, but its depth would have depended on the presence of a nuchal notch. The remaining 10 pairs of peripherals form a continuous ring that surrounds the central carapacial disk.
Carapacial scutes
The carapace of Peltochelys duchastelii is covered by 5 vertebrals, 4 pairs of pleurals, and at least 11 pairs of marginals (Fig. 1). The anterior margin of the shell is damaged, obscuring the likely presence of another pair of marginals and perhaps also a cervical. To maintain homology with other turtles, we label the most anterior preserved marginal as marginal 2, which results in a count of 12 pairs of marginals. The interpretation is supported by external observations. In particular, the interpleural sulci of the pleurals laterally contact marginals 5, 7, and 9, as in all other turtles, including those with 11 pairs of marginals. The alternative interpretation, as suggested but not explicitly discussed by Meylan (1988) or Pérez-García (2011), in contrast, suggests highly apomorphic contacts of the interpleural sulci with marginals 4, 6, and 8.
Vertebral 1 is the broadest vertebral element and therefore clearly covers the median portions of peripheral 2 (Fig. 1). It has a broad lateral contact with marginal 2, a broad posterolateral contact with pleural 1, a broad posterior contact with vertebral 2, and an unusual lateral point contact with marginal 3. Possible contacts with marginal 1 and the cervical, if present, are obscured by damage. Vertebrals 2–4 have irregular hexagonal outlines created by their sinuous lateral margins with the pleurals. They are narrower than vertebral 1 and decrease in width from anterior to posterior. Vertebral 5 is a heptagonal element that is about as wide as vertebral 2. It has an anterolateral contact with pleural 4 and similarly sized posterior contacts with marginals 11 and 12. In our counting system, the intervertebral sulci are located at neurals 1, 3, and 6, and suprapygal 1 and the interpleural sulci are located as costals 2, 4, 6, and 8. As suggested above, the interpleural sulci run laterally into marginals 5, 7, and 9. The marginals are narrow elements that do not cover the costals or suprapygals.
Plastral bones
There is full agreement that Peltochelys duchastelii lacks mesoplastra (Fig. 1). The plastron therefore consists of an entoplastron and paired epiplastra, hyoplastra, hypoplastra, and xiphiplastra. The anterior plastral lobe has a straight anterior margin and covers much of the anterior carapacial opening. The posterior plastral lobe is narrower than the anterior plastral lobe and only partially covers the posterior carapacial opening. A rounded, shallow anal notch is present. Neither lobe shows signs of kinesis.
The buttresses are visible on the left side of the lectotype, but disarticulation and crushing obscure their contacts (Fig. 1). We therefore conclude that the axillary buttresses are well developed and likely contacted the costals but also that the contacts with the peripherals are unclear. A lack of a sutural articulation site at peripheral 8 further suggests that the inguinal buttress only reached peripheral 7, but it is unclear if it reached the overlying costals as well. Previous authors did not discuss the contacts of the bridge explicitly.
Plastral scutes
There is disagreement in the number of plastral scales for Peltochelys duchastelii. All authors agree that this turtle possesses paired extragulars, humerals, pectorals, abdominals, femorals, and anals, but while Hummel (1929) and Meylan (1988) suggest the presence of a single, median gular, Pérez-García (2011) reconstructs the presence of highly asymmetric, paired gulars. In our opinion, the gular–gular sulcus highlighted by Pérez-García (2011) resembles one of many cracks that run through the anterior plastral lobe. We therefore side here with Hummel (1929) and Meylan (1988). Our observations otherwise fully agree with those of previous authors: the midline sulcus is sinuous along its full length, the extragulars are triangular elements that do not cover the entoplastron and hyoplastron, the gular is a broad element that laps onto the anterior tip of the entoplastron, the humeral–pectoral sulcus is located far behind the entoplastron, the abdominal contributes to the inguinal notch, and the anals are clearly restricted to the xiphiplastra (Fig. 1). At least three pairs of inframarginals are present, of which the first contacts the pectoral, the second covers the hyoplastral–hypoplastral suture and contacts the pectoral and abdominal, and the third contacts the abdominal.
To investigate the phylogenetic relationships of Peltochelys duchastelii, we inserted it into modifications of the three analyses employed by Lyson and Joyce (2011) to investigate the placement of Compsemys victa.
The first analysis, which is based on the matrix of Gaffney et al. (2007), was expanded to include Peltochelys duchastelii (see Supplement) and then subjected to a parsimony analysis using TNT (Goloboff et al., 2008) with standard protocols (i.e., 1000 random addition sequences followed by a round of tree bisection reconnection). The composite terminal taxon “Synapsida/Diapsida” was maintained as the outgroup. A light implied weight with a k value of 12 was employed following the recommendations of Goloboff et al. (2018). A summary of the resulting single most parsimonious tree with a length of 7.04066 is provided in Fig. 2a.
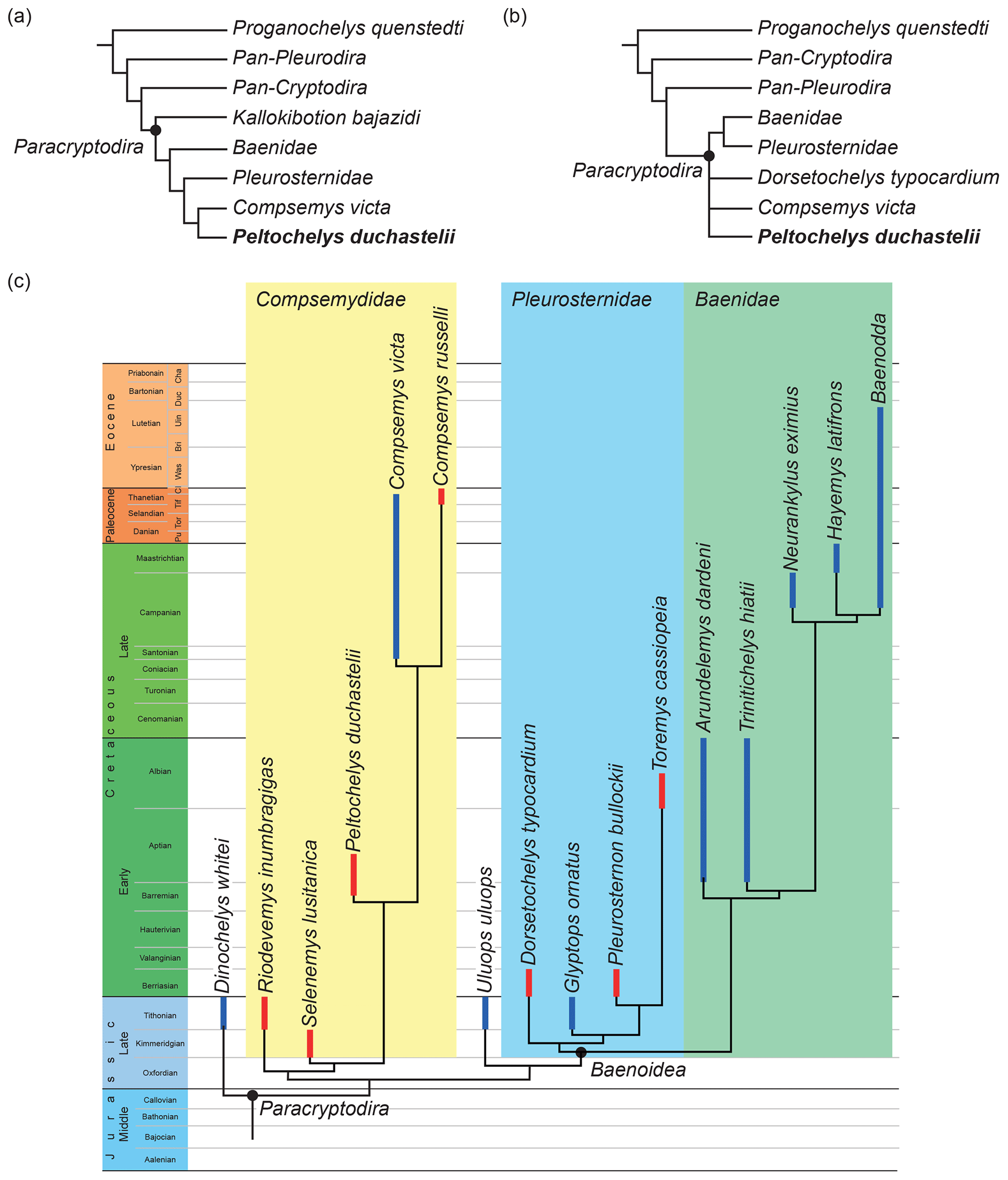
Figure 2The results of three separate phylogenetic hypotheses that include Peltochelys duchastelii. (a) The single most parsimonious tree with a length of 7.04066 resulting from the expanded analysis of Gaffney et al. (2007). (b) The strict consensus of the 27 most parsimonious solutions with a length of 13.43147 resulting from the updated analysis of Joyce (2007). (c) A time-calibrated extract of the single most parsimonious solution with a length of 9.22411 resulting from the expanded analysis of Lyson and Joyce (2011). The range of North American taxa is highlighted in blue and that of European taxa is highlighted in red.
The second analysis, which is based on the matrix of Joyce (2007), already included Peltochelys duchastelii, but its scoring was corrected following the observations made herein (see Supplement). These changes mostly overlap with those implemented by Pérez-García (2011) for the same matrix. Two new characters were added to the analysis (137, peripheral 1 and costal 1 contact, 0 = present, 1 = absent; 138, contribution of nuchal to margin of shell, 0 = present, 1 = absent). The same settings were used for the parsimony analysis as for the first analysis, but 15 characters were ordered (7, 27, 33, 35, 54, 60, 61, 65, 68, 71, 85, 98, 120, 133, and 134). The hypothetical ancestor was maintained as the outgroup. A summary of the strict consensus of the 27 most parsimonious solutions with a length of 13.43147 is provided in Fig. 2b.
As the first two analysis suggest paracryptodire relationships for Peltochelys duchastelii in a global context, we added this taxon to the paracryptodire matrix of Lyson and Joyce (2011). In addition to Peltochelys duchastelii, the matrix was expanded to include Compsemys (Berruchelus) russelli from the Paleocene of France, as described by Pérez-García (2012); Riodevemys inumbragigas from the Early Cretaceous of Spain, as described by Pérez-Garcia et al. (2015a); Selenemys lusitanica from the Late Jurassic of Portugal, as described by Pérez-García and Ortega (2011); and Toremys cassiopeia from the Early Cretaceous of Spain, as described by Pérez-Garcia et al. (2015b). The scoring of Dorsetochelys typocardium was updated following Pérez-Garcia (2014). Characters 42, 43, 49, and 51 (formerly 52) were expanded to consist of three character states that can be ordered (character 42: 0 = mesoplastra present with midline contact; 1 = mesoplastra present, but midline contact absent; 2 = mesoplastra absent; character 43: 0 = cervicals absent; 1 = one cervical present; 2 = two or more cervicals present; character 49: 0 = the anterior margin of marginal 1 mostly covers the nuchal; 1 = the anterior margin of marginal 1 evenly straddles the nuchal and peripheral 1 suture; 2 = the anterior margin of marginal 1 mostly covers peripheral 1; character 51: gulars much smaller than extragulars; 1 = gulars similar in size to extragulars; 2 = gulars much larger than extragulars). Eight characters were omitted from the analysis, as they were originally formulated to resolve relationships deep within Baenidae and are inapplicable to the base of Paracryptodira (characters 50, 55, 59, 60, 100, 101, 103, and 105 of Lyson and Joyce, 2011). Many of these changes resemble those implemented by Pérez-García (2012) and Pérez-García et al. (2015a, b) for the same matrix, but numerous differences remain. Our matrix is therefore a somewhat independent, though related, analysis to those of Pérez-García et al. (2015a, b). The final matrix (see Supplement) was subjected to a parsimony analysis using the same settings as the first two analyses, including implied weighting, but 15 characters were run ordered (6, 14, 16, 18, 28, 33, 36, 42, 43, 44, 49, 51, 63, 66, and 101). Proganochelys quenstedtii was selected as the outgroup. An extract of the single most parsimonious solution with a length of 9.22411 is provided in Fig. 2c.
4.1 Morphology
Our interpretation of the shell of the lectotype of Peltochelys duchastelii differs substantially from the recent ones provided by Meylan (1988) and Pérez-García (2011), most importantly in regards to the nuchal, the peripheral series, and the marginals. Unfortunately, these differences are mostly a result of the preservation of this specimen because it is flattened, many bones are fractured and have shifted slightly from their original positions, and it was painted in the 19th century to highlight the sulci and sutures in the photographs that accompany the descriptions of Dollo (1884). As we cannot discount the possibility that our observations are erroneous, we here highlight several lines of evidence that provide external support for our interpretation. We order these from the strongest to the weakest.
-
To our knowledge, costal rib 1 inserts laterally into peripheral 3 in all crown turtles with a generalized shell, even in those with 10 peripherals. In the interpretation of Meylan (1988) and Pérez-García (2011), Peltochelys duchastelii would be the lone exception to this rule in that costal rib 1 would insert laterally into peripheral 2. In our interpretation, by contrast, Peltochelys duchastelii is consistent with all other turtles by having costal rib 1 insert laterally into peripheral 3.
-
To our knowledge, the interpleural sulci of all crown turtles with a generalized shell insert distally into marginals 5, 7, 9, even in those turtles that only have 10 pairs of marginals. The alternative interpretations of Meylan (1988) and Pérez-García (2011), by contrast, suggest the apomorphic contact of the interpleural sulci with marginals 4, 6, and 8. Although we cannot observe marginal 1 in the lectotype of Peltochelys duchastelii, as the relevant portion of the shell is not preserved, the presence of 11 peripherals calls for the presence of 12 marginals, of which marginals 5, 7, and 9 contact the interpleural sulci.
-
The bones of the carapace of most turtles usually contact one another along straight sutures, of which three typically converge at obtuse angles upon points which resemble triple junctions formed by tectonic plates. As a result, most bones can be conceptualized as non-convex polygons. In the interpretation of Meylan (1988) and Pérez-García (2011), the nuchal is laterally notched to hold what they interpret as the first peripheral, a morphology not seen in any other turtle. In our interpretation, by contrast, peripheral 1 forms a straight contact with peripheral 2 and peripheral 2 forms a straight contact with the nuchal. As a result, the nuchal is a non-convex polygon in our interpretation.
-
Our interpretation is made plausible by the existence of similar arrangements to the nuchal and anterior peripherals in the broadly coeval paracryptodires Compsemys (Berruchelus) russelli (Pérez-García, 2012), Compsemys victa (Gilmore, 1919), Selenemys lusitanica (Pérez-Garcia and Ortega, 2011), Toremys cassiopeia (Pérez-Garcia et al., 2015a), and the taxonomically problematic Kallokibotion bajazidi (Pérez-García and Codrea, 2018). We therefore are not suggesting a unique morphology, just one that differs from previous interpretations.
-
The observation that peripherals 1 contact one another anterior to the nuchal was independently reported by Meylan (1988) for the now lost juvenile from Bernissart. This suggests that our interpretation is not foreign to turtles from this locality.
-
Our interpretation overlaps with that of the person, likely Dollo (1884), who painted the specimen towards the end of the 19th century. This is relevant, as this person was the last to see the specimen with uncovered sutures.
-
Last, but not least, Dollo (1884) highlighted that the necks of the juveniles from Bernissart show a lateral bend more consistent with pleurodires. Lateral head motion is also consistent with paracryptodires (Werneburg et al., 2015) but not with crown cryptodires, in particular pan-trionychians, which show a hyper-specialized cryptodiran neck motion (Meylan and Gaffney, 1989).
In conclusion, although we cannot guarantee that our interpretation is without errors, numerous lines of evidence highlight that our hypothesis is highly plausible. The ultimate test, however, will be the discovery of new, better preserved material, which independently weigh in favor of one over the other hypothesis.
4.2 Alpha taxonomy
The Early Cretaceous locality of Bernissart has yielded a total of five fossil turtles that we here refer to Peltochelys duchastelli. The shell of the lectotype (Fig. 1) overall resembles that of many other river and pond turtles, notably paracryptodires, pleurodires, adocids, and testudinoids, by being oval and having broad plastral lobes with well-developed axillary buttress. A unique combination of characters, in particular the retracted nuchal combined with a lack of mesoplastra, clearly distinguishes this taxon from all other known turtles. The validity of this species is therefore unproblematic. Dollo (1884) and Pérez-Garcia (2015) reported four juvenile specimens from Bernissart that only display a few anatomical features. However, as the available morphology is consistent with Peltochelys duchastelii, we here agree with Pérez-Garcia (2015) that all four can be referred to this species using spatial and temporal considerations. Meylan (1988) reported the presence of a fifth juvenile from Bernissart but concluded that it is not referable to Peltochelys duchastelii as the peripherals excluded the nuchal from the anterior margin of the shell. Although this specimen was never figured and has since been reported to be lost (Pérez-García, 2011, 2015), the limited character evidence supports referral to Peltochelys duchastelii after all, while confirming the veracity of the observation we make for the lectotype. We therefore agree here with Pérez-García (2014) and Pérez-García et al. (2015b) that Peltochelys duchastelii is restricted to Bernissart for the moment.
4.3 Phylogenetic relationships
Over the course of the last century, Peltochelys duchastelii has been argued to be a pleurodire (Dollo, 1884), a chelydrid (Lydekker and Boulenger, 1887), a dermatemydid (Nopcsa, 1928a, b), a stem carettochelyid (Hummel, 1929, 1932; Lapparent de Broin, 2001), an adocusian (Chkhikvadze, 1975; Nessov, 1977), or a stem trionychian (Meylan, 1988; Brinkman, 1998; Meylan and Gaffney, 1989; Brinkman and Peng, 1996; Joyce, 2007; Perez-García, 2011) but has never been associated with paracryptodires. This is somewhat surprising, as Cretaceous sediments in Europe are dominated by paracryptodires (Joyce and Anquetin, 2019), and the textured shell of Peltochelys duchastelii globally resembles that of other paracryptodires. We are unable to reconstruct the thought process of past researchers, as none discussed a possible link with paracryptodires explicitly or extensively, but we speculate upon an overreliance on “key characters”: in this case the absence of mesoplastra. These bones symplesiomorphically occur at the base of Testudinata but were lost independently at least three times, in particular along the stem lineages of Chelidae and Cryptodira (Gaffney et al., 1991; Joyce, 2007) but also within Meiolaniformes (Sterli et al., 2015). The consistent presence or absence of mesoplastra in most groups of turtles renders this bone a useful initial guide in establishing phylogenetic relationships, but an overreliance on this character appears to have misled researchers in the past.
To assess the phylogenetic relationships of Peltochelys duchastelii, we followed the protocols of Lyson and Joyce (2011) for their analysis of Compsemys victa by first inserting this turtle into the global turtle matrices of Gaffney et al. (2007) and Joyce (2007). In both cases, the analysis retrieved Peltochelys duchastelii as the sister to Compsemys victa within Paracryptodira, mostly based on the presence of a sinuous plastral midline sulcus, a sutured bridge, absence of a contact between peripheral 1 and costal 1, and the withdrawal of the nuchal from the anterior margin of the shell. As we find the listed similarities with Compsemys victa, an uncontroversial paracryptodire, to be of particular relevance, we further expanded the paracryptodire analysis of Lyson and Joyce (2011) to all named Cretaceous paracryptodires from Europe in addition to Peltochelys duchastelii. The resulting matrix is similar but not identical to that of Pérez-García et al. (2015a, b). The analysis nevertheless retrieves a tree that greatly differs from that of Pérez-García et al. (2015a, b), by recognizing a basal clade of mostly European paracryptodires consisting of the Late Jurassic Selenemys lusitanica and Riodevemys inumbragigas, the Early Cretaceous Peltochelys duchastelii, and the Late Cretaceous to Paleogene Compsemys russelli and Compsemys victa. The full clade is diagnosed by the presence of a sinuous plastral midline sulcus. The clade that excludes Riodevemys inumbragigas is united by the absence of cervicals, the related expansion of marginal 1 onto the nuchal, a midline contact of peripherals 1 resulting in a withdrawal of the nuchal from the margin of the shell, and loss of a contact between peripheral 1 and costal 1. The clade consisting of Peltochelys duchastelii and both species of Compsemys is united by square vertebral scutes. Peltochelys duchastelii is excluded from the Compsemys clade by lacking enlarged gulars and a deep xiphiplastral notch. The placement of Peltochelys duchastelii within Paracryptodira is therefore based on a series of characters that are otherwise unknown in turtles, in particular the arrangement of the bones and scutes in the nuchal area. Naturally, all of our conclusions are based on our novel interpretation of the lectotype specimen.
4.4 Peltochelyidae versus Compsemydidae
Two names exist that could be applied to the clade that includes Compsemys: Peltochelyidae and Compsemydidae. The term Peltochelyidae was initially created by Seeley (1880) to unite all turtles with a “granular surface structure”, in particular Trionychidae. As Peltochelys was not named until a few years later (Dollo, 1884), Seeley (1880) did not create an available name, as the ICZN (1999) demands that a new family name created prior to 1931 be at least based on an available genus name. The conclusion that Seeley (1880) is the author of Peltochelyidae, as first suggested by Lapparent de Broin (2001), is therefore not correct. Boulenger (1889) listed Peltochelyidae as a synonym of Pleurodira and Trionychidae, and Hay (1902) listed it as a synonym of Trionychia. As both authors did not believe this taxon to be valid, these authors did not make this name available as well, as the ICZN (1999) demands that all new names be used as valid to be made available. The name Peltochelyidae is used for the first time as valid for the group of turtles that includes Peltochelys duchastelii by Lapparent de Broin (2001), followed, among others, by Danilov (2005) and Karl et al. (2007, 2012). To avoid the unintentional naming of new family names, the ICZN (1999) demands that family names created after 1999 are explicitly highlighted as such. As none of these newer contributions suggest the creation of a new family name, Peltochelyidae remains unavailable to date. The term Compsemyidae was introduced by Zangerl (1969), likely for Compsemys victa, but as the term is neither associated with a definition nor an indication, it cannot be considered available. The name Compsemydidae, by contrast, was explicitly introduced as a new family name by Pérez-García et al. (2015b). We therefore here informally convert this name to the most inclusive group of turtles that includes Compsemys victa but not the baenid Baena arenosa Leidy, 1870, the pleurosternid Pleurosternon bullockii (Owen, 1842), or any extant turtle.
4.5 Biogeographic considerations
In addition to the strong character evidence listed above, the newly recognized compsemydid clade that includes Peltochelys duchastelii is also supported by temporal and biogeographic considerations, as it mostly unites continental turtles from the Late Jurassic to Paleogene of western Europe and suggests that Peltochelys duchastelii originated in situ. The sole exception to the rule is Compsemys victa, which is known from Campanian to Paleogene sediments across Laramidia (Lyson and Joyce, 2011). As Compsemys victa had initially been reported from North America, recent biogeographic models suggested that it is indigenous to this continent but immigrated to Europe using the Thulean route, together with many other faunal elements (Brikiatis, 2014), following the K–T extinction event, to give rise to Compsemys russelli (Pérez-García, 2012). The most parsimonious interpretation of our cladogram, by contrast, suggests that the Compsemys lineage is indigenous to Europe instead and that the ancestor of Compsemys victa immigrated from there to North America prior to the Campanian. The proposed migration, however, is inconsistent with the movement of other taxa at the time, and suggests overcoming multiple barriers, in particular the North Atlantic and the Western Interior Seaway.
A less parsimonious possibility remains that is nevertheless broadly consistent with the fossil record. It is possible that the immediate Compsemys lineage extends to the Jurassic of Euramerica, only survived in North America following the opening of the North Atlantic, and then immigrated to Europe following the K–T boundary event, as previously proposed. This interpretation is supported by previous insights into the vicariance of paracryptodires (Hirayama et al., 2000; Joyce et al., 2016), commonly used dispersal routes (Brikiatis, 2014; Joyce et al., 2016), and the presence of Late Jurassic compsemydids, such as Selenemys lusitanica.
The character/taxon matrices using in this study are available in the Supplement.
The supplement related to this article is available online at: https://doi.org/10.5194/fr-23-83-2020-supplement.
WGJ and YR photographed, illustrated, and described the material; performed the phylogenetic analysis; and wrote the primary draft of the manuscript. All authors read and approved the final draft.
The authors declare that they have no conflict of interest.
We would like to thank Annelise Folie for providing access to specimens in her care. Igor Danilov, Jordan Gônet, and Adán Pérez-García provided numerous thoughtful comments that greatly helped improve the quality of this paper. This study was supported by a grant from the Swiss National Science Foundation to WGJ (SNF 200021_178780/1).
This research has been supported by the Swiss National Science Foundation (grant no. SNF 200021_178780/1).
This paper was edited by Torsten Scheyer and reviewed by Adán Pérez-García, Igor Danilov, and Jordan Gônet.
Batsch, A. J. G. C.: Versuch einer Anleitung, zur Kenntniß und Geschichte der Thiere und Mineralien, Akademische Buchhandlung, Jena, Germany, 1788.
Boulenger, G. A.: Catalogue of Chelonians, Rhynchocephalians and Crocodiles in the British Museum (Natural History), Taylor and Francis, London, UK, 1889.
Brikiatis, L.: The De Geer, Thulean and Beringia routes: key concepts for understanding early Cenozoic biogeography, J. Biogeogr., 41, 1036–1054, 2014.
Brinkman, D.: The skull and neck of the Cretaceous turtle Basilemys (Trionychoidea, Nanhsiungchelyidae), and the interrelationships of the genus, Paludicola, 1, 150–157, 1998.
Brinkman, D. and Peng, J.-H.: A new species of Zangerlia (Testudines: Nanhsiungchelyidae) from the Upper Cretaceous redbeds and Bayan Mandahu, Inner Mongolia, and the relationships of the genus, Can. J. Earth Sci., 33, 526–540, 1996.
Broin, F. de: Contribution à l'étude des Chéloniens. Chéloniens continentaux du Crétacé et du Tertiaire de France, Mem., Mus. Nat. Hist. Nat., C38, 1–366, 1977.
Danilov, I. G.: Die fossilen Schildkröten Europas, in: Handbuch der Reptilien und Amphibien Europas; Part 3/IIIB: Schildkröten (Testudines) II, edited by: Fritz, U., AULA-Verlag, Wiebelsheim, Germany, 329–448, 2005.
Danilov, I. G., Syromyatnikova, E. V., and Sukhanov, V. B.: Subclass Testudinata, in: Fossil vertebrates of Russia and adjacent countries. Fossil Reptiles and Birds. Part 4, edited by: Lopatin, A. V. and Zelenkov, N. V., GEOS, Moscow, Russia, 27–395, 2017 (in Russian).
Dollo, L.: Première note sur les chéloniens de Bernissart, Bull. Mus. R. Hist. Nat. Belg., 3, 63–79, 1884.
Dollo, L.: The fossil vertebrates of Belgium, Ann. N.Y. Acad. Sci., 19, 99–119, 1909.
Chkhikvadze, V. M.: Volume and systematic position of turtles of the suborder Amphichelydia Lydekker, 1889, Soob. Akad. Nauk Gruzinskoy SSR, 78, 745–748, 1975 (in Russian).
Gaffney, E. S.: A phylogeny and classification of higher categories of turtles, B. Am. Mus. Nat. Hist., 155, 387–436, 1975.
Gaffney, E. S., Meylan, P. A., and Wyss, A. R.: A computer assisted analysis of the relationships of the higher categories of turtles, Cladistics, 7, 313–335, 1991.
Gaffney, E. S., Rich, T. H., Vickers-Rich, P., Constantine, A., Vacca, P., and Kool, L.: Chubutemys, a new eucryptodiran turtle from the Early Cretaceous of Argentina, and the relationships of the Meiolaniidae, Am. Mus. Novit., 3599, 1–35, 2007.
Gilmore, C. W.: New fossil turtles, with notes on two described species, Proc. U. S. Nat. Mus., 56, 113–132, 1919.
Goloboff, P. A., Farris, J., and Nixon, K.: TNT: a free program for phylogenetic analysis, Cladistics, 24, 774–786, 2008.
Goloboff, P. A., Torres, A., and Salvador Arias, J.: Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology, Cladistics, 34, 407–437, 2018.
Hay, O. P.: Bibliography and catalogue of the fossil vertebrata of North America, Bull. U. S. Geol. Surv., 179, 1–868, 1902.
Hirayama, R., Brinkman, D. B., and Danilov, I. G.: Distribution and biogeography of non-marine Cretaceous turtles, Russ. J. Herpetol., 7, 181–198, 2000.
Hummel, K.: Die fossilen Weichschildkröten (Trionychia). Ein morphologisch-systematische und stammesgeschichtliche Studie, Geol. Pal. Abh., 16, 359–487, 1929.
Hummel, K.: Fossilium Catalogus. I: Animalia. Pars 52: Trionychia fossilia, W. Junk, Berlin, Germany, 1932.
ICZN (International Commission of Zoological Nomenclature): International Code of Zoological Nomenclature, 4th Edn., International Trust for Zoological Nomenclature, London, UK, 1999.
Joyce, W. G.: Phylogenetic relationships of Mesozoic turtles, B. Peabody Mus. Nat. Hi., 48, 3–102, 2007.
Joyce, W. G.: A review of the fossil record of basal Mesozoic turtles, B. Peabody Mus. Nat. Hi., 58, 65–113, 2017.
Joyce, W. G. and Anquetin, J.: A review of the fossil record of nonbaenid turtles of the clade Paracryptodira, B. Peabody Mus. Nat. Hi., 60, 129–155, 2019.
Joyce, W. G., Sterli, J., and Chapman, S. D.: The skeletal morphology of the solemydid turtle Naomichelys speciosa from the Early Cretaceous of Texas, J. Paleontol., 88, 1257–1287, 2014.
Joyce, W. G., Rabi, M., Clark, J. M., and Xu, X.: A toothed turtle from the Late Jurassic of China and the global biogeographic history of turtles, BMC Evol. Biol., 16, 236, https://doi.org/10.1186/s12862-016-0762-5, 2016.
Karl, H.-V., Staesche, U., Tichy, G., Lehmann, J., and Peitz, S.: Systematik der Schildkröten (Anapsida: Chelonii) aus Oberjura und Unterkreide von Nordwestdeutschland, Geol. Jb., B98, 5–89, 2007.
Karl, H.-V., Milan, J., and Tichy, G.: New shell remains of Peltochelys duchastelli (Dollo, 1884) from western and middle Europe and the rise of Trionychids (Testudines: Trionychoidea), Stud. Geol. Salmant., 9, 87–106, 2012.
Lapparent de Broin, F. de: The European turtle fauna from the Triassic to the Present, Dumerilia, 4, 155–217, 2001.
Leidy, J.: Descriptions of Emys jeanesi, Emys haydeni, Baena arenosa, and Saniwa ensidens, P. Acad. Nat. Sci. Phila., 1870, 123–124, 1870.
Lydekker, R.: Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History). Part III, The Order Chelonia, Trustees of the British Museum, London, UK, 1889.
Lydekker, R. and Boulenger, G. A.: Notes on Chelonia from the Purbeck, Wealden and London-clay, Geol. Mag., 4, 270–275, 1887.
Lyson, T. R. and Joyce, W. G.: Cranial anatomy and phylogenetic placement of the enigmatic turtle Compsemys victa Leidy, 1856, J. Paleontol., 85, 789–801, 2011.
Mantell, G. A.: The Geology of the South East of England, Longman, London, UK, 1833.
Meylan, P. A.: Peltochelys Dollo and the relationships among the genera of the Carettochelyidae (Testudines, Reptilia), Herpetologica, 44, 440–450, 1988.
Meylan, P. A. and Gaffney, E. S.: The skeletal morphology of the Cretaceous cryptodiran turtle, Adocus, and the relationships of the Trionychoidea, Am. Mus. Novit., 2941, 1–60, 1989.
Młynarski, M.: Handbuch der Paläoherpetologie. Part 7, Testudines. Gustav Fischer Verlag, Stuttgart, Germany, 1976.
Nessov, L. A.: Skull morphology of Early Cretaceous turtles of the family Adocidae, Trud. Zool. Inst. AN SSSR, 74, 75–80, 1977 (in Russian).
Nopcsa, F.: Palaeontological notes on reptiles, Geol. Hung. Palaeontol Ser., 1, 1–84, 1928a.
Nopcsa, F.: The genera of reptiles, Palaeobiol., 1, 163–188, 1928b.
Owen, R.: Report on British fossil reptiles. Part II, Rep. Brit. Ass. Adv. Sci., 11, 60–204, 1842.
Pérez-García, A.: Revisión de la serie tipo de Peltochelys duchastelii (Chelonii, Trionychoidea) del Cretácico Inferior de Bernissart (Bélgica), in: Viajando a Mundos Pretéritos, edited by: Pérez-García, A., Gascó, F., Gasulla, J. M., and Escaso, F., Ayuntamiento de Morella, Castellón, Spain, 249–260, 2011.
Pérez-García, A.: Berruchelus russelli gen. et sp. nov., a paracryptodiran turtle from the Cenozoic of Europe, J. Vertebr. Paleontol., 32, 545–556, 2012.
Pérez-García, A.: Revision of the poorly known Dorsetochelys typocardium, a relatively abundant pleurosternid turtle (Paracryptodira) in the Early Cretaceous of Europe, Cret. Res., 49, 152–162, 2014.
Pérez-García, A.: Dos ejemplares inéditos de tortugas hallados en el siglo XIX en el Cretácico Inferior de Bernissart (Bélgica), Geogaceta, 57, 67–70, 2015.
Pérez-García, A. and Codrea, V.: New insights on the anatomy and systematics of Kallokibotion Nopcsa, 1923, the enigmatic uppermost Cretaceous basal turtle (stem Testudines) from Transylvania, Zool. J. Linn. Soc.-Lond., 182, 419–443, 2018.
Pérez-García, A. and Ortega, F.: Selenemys lusitanica, gen. et sp. nov., a new pleurosternid turtle (Testudines: Paracryptodira) from the Upper Jurassic of Portugal, J. Vertebr. Paleontol., 31, 60–69, 2011.
Pérez-García, A., Espílez, E., Mampel, L., and Alcalá, L.: A new European Albian turtle that extends the known stratigraphic range of the Pleurosternidae (Paracryptodira), Cret. Res., 55, 74–83, 2015a.
Pérez-García, A., Royo-Torres, R., and Cobos, A.: A new European Late Jurassic pleurosternid (Testudines, Paracryptodira) and a new hypothesis of paracryptodiran phylogeny, J. Syst. Palaeontol., 13, 351–369, 2015b.
Seeley, H. G.: Note on Psephophorus polygonus, v. Meyer, a new type of Chelonian reptile allied to the leathery turtle, Q. J. Geol. Soc. Lond., 36, 406–413, 1880.
Sterli, J., de la Fuente, M. S., and Umazano, A. M.: New remains and new insights on the Gondwanan meiolaniform turtle Chubutemys copelloi from the Lower Cretaceous of Patagonia, Argentina, Gond. Res., 27, 978–994, 2015.
Wegner, T.: Desmemys Bertelsmanni n. g. n. sp. Ein Beitrag zur Kenntnis der Thalassemydidae Rütimeyer, Palaeontogr., 58, 105–132, 1911.
Werneburg, I., Hinz, J., Gumpenberger, M., Volpato, V., Natchev, N., and Joyce, W. G.: Modeling neck mobility in fossil turtles, J. Exp. Zool. Part B, 324, 230–243, 2015.
Yans, J., Dejax, J., Pons, D., Dupuis, Ch., and Taquet, P.: Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique) (Palaeontological and geodynamical implications of the palynological dating of the wealden facies sediments of Bernissart [Mons Basin, Belgium]), C.R. Palévol, 4, 135–150, 2005.
Zangerl, R.: The turtle shell, in: The Biology of the Reptilia, Vol. 1, edited by: Gans, C. and Parsons, T. S., Academic Press, London, 311–339, 1969.