the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 26 Jul 2021
| 26 Jul 2021
At the crossroads: early Miocene marine fishes of the proto-Mediterranean Sea
Efterpi Koskeridou
Danae Thivaiou
Connectivity and climate control fish distribution today as well as in the geological past. We present here the Aquitanian (early Miocene) marine fish of the Mesohellenic Basin, a restricted basin at the border between the proto-Mediterranean and Paratethyan seas. Based on fish otoliths, we were able to identify 19 species from 17 genera, including two new species: Ariosoma mesohellenica and Gnathophis elongatus. This fish assemblage, in conjunction with the accompanying molluscan assemblage, indicates a variable shelf paleoenvironment with easy access to the open ocean. Although available data on the Indo-Pacific fishes of the early Miocene are very limited, the fish fauna of the Mesohellenic Basin has many elements in common with the North Sea, the NE Atlantic, and the Paratethys.
- Article
(3913 KB) -
Supplement
(1144 KB) - BibTeX
- EndNote





Connectivity between marine realms largely controls the fate of fish populations and biodiversity (Worm and Tittensor, 2018). Early Miocene proto-Mediterranean biodiversity is particularly interesting because, at that time, the basin's connection with the Indo-Pacific realm ceased to exist (Rögl, 1998; Bialik et al., 2019; Torfstein and Steinberg, 2020; Sun et al., 2021), while the marine biodiversity hotspot was shifting toward the southeast (Renema et al., 2008) and the climate was cooling (Cornacchia et al., 2018), marking the onset of a subtropical gradually oligotrophic regime in the proto-Mediterranean Sea. However, the early Miocene fossil fish record from the Mediterranean area is particularly sparse. Moreover, very few studies have focused on the fish otolith record (Steurbaut, 1979, 1982, 1984; Reichenbacher and Cappetta, 1999; Reichenbacher, 2004; Hoedemakers and Batllori, 2005), despite its importance for revealing the evolution of fish faunas and fish biogeography (Agiadi et al., 2011, 2017, 2018). At the crossroads between the proto-Mediterranean Sea, the Atlantic Ocean, the North Sea, the Paratethys, and the Indo-Pacific realm, the Mesohellenic Basin (MHB) during the early Miocene, a molassic basin at the northern part of the proto-Mediterranean, directly at the intersection with the Paratethys epicontinental sea, offers a unique opportunity to explore the effects of interbasinal connectivity on fish faunas.
In this study, we identify the fossil fish otoliths found in the Aquitanian (lower Miocene) sediments of the Felli section (Grevena Prefecture, NW Greece) that were deposited in the MHB. The reconstructed assemblage is the first record of fishes from the eastern part of the Aquitanian (∼ 23–20 Ma) proto-Mediterranean region. Furthermore, we discuss the paleobiogeographic range of the identified fish species in order to draw a conclusion regarding the factors that may have determined the subsequent evolution of the Mediterranean marine fish fauna.
The study area is located in Grevena Prefecture, NW Greece, where the molassic sedimentary sequence of the MHB is exposed (Fig. 1; Zelilidis et al., 2002; Ferrière et al., 2004). The MHB was a synorogenic molasse-type basin (Kilias et al., 2015), a separate paleogeographic area from the late Eocene until the middle Miocene, between the internal and external Hellenides, ENE and WSW, respectively. It was a narrow, 300 km long trench, filled with up to 4.5 km of sediments, which have been separated into the Krania, Eptachori, Pentalofos, Tsotyli, and Ondria formations (Brunn, 1956; Wielandt-Schuster et al., 2004; Ferrière et al., 2013; Kilias et al., 2015), deposited parallel to each other (except for the Krania Formation) along the eastern margin of the trough (Kilias et al., 2015). The studied sediments belong to the Pentalofos Formation, consisting in general of upper Chattian–lower Miocene sandstones to silty marlstones (Mavridis et al., 1985; Ferrière et al., 2004; Wielandt-Schuster et al., 2004; Kilias et al., 2015). The Felli section, which is our study section, includes only the uppermost part of the Pentalofos Formation corresponding to the Aquitanian according to Mavridis et al. (1985). The Pentalofos Formation overlies the Eptachori Formation and is overlain, conformably in the Grevena area, by the Tsotyli Formation (Fermeli and Ioakim, 1992).
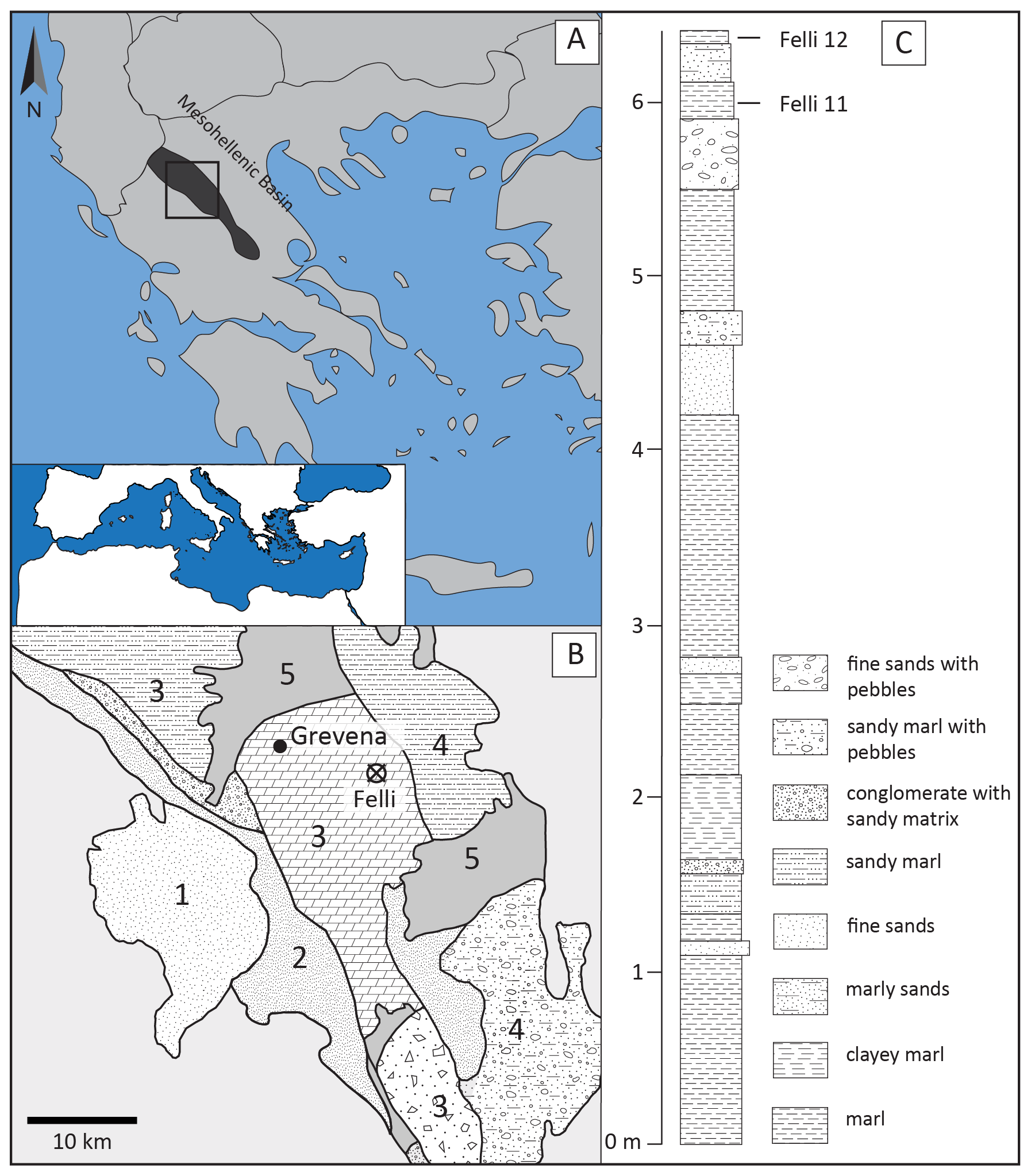
Figure 1(A) Map of Greece where the Mesohellenic Basin is indicated. (B) Geological map of the study area: (1) Krania Formation (Eocene), (2) Eptachori Formation (lower–upper Oligocene), (3) Pentalofos Formation (upper Oligocene–lower Miocene), (4) Tsotyli Formation (middle Miocene), and (5) Quaternary. (C) Lithostratigraphic column of the Felli section with the sampled levels (modified after Zelilidis et al., 2002, and Thivaiou et al., 2019).
We sampled a small section by the Aliakmon River banks, located east of Felli village, SE of Grevena City (40∘01′4.55′′ N, 21∘33′34.37′′ E). The section is mainly composed of conglomerates and sands, and the grain size decreases toward the top, where we find marls and then clays. We obtained ∼ 20–25 kg of sediment from each of two marl–clay beds, F11 and F12, following the numbering scheme of Thivaiou et al. (2019). The samples were water-sieved using a 250 µm sieve, and the residues were dried in an oven. The otoliths were handpicked from the residues and identified under the microscope.
We adopted specific descriptive characters for the otoliths of Congridae (Schwarzhans, 2019) and Pleuronectiformes (Schwarzhans, 1999). We followed the fish classification scheme of Nelson et al. (2016). We made the following measurements on the identified otoliths (reported in mm; Fig. 2): OL, otolith length; OH, otolith height; AL, antirostrum length; RL, rostrum length; CL, colliculum length; OCL, ostial colliculum length; CCL, caudal colliculum length; OSL, ostium length; CaL, caudal length. In addition, in Congridae, it proved significant to measure the sulcus angle (α), as the angle formed between the sulcus direction and the otolith length direction (Fig. 2). All data can be found in the Supplement.
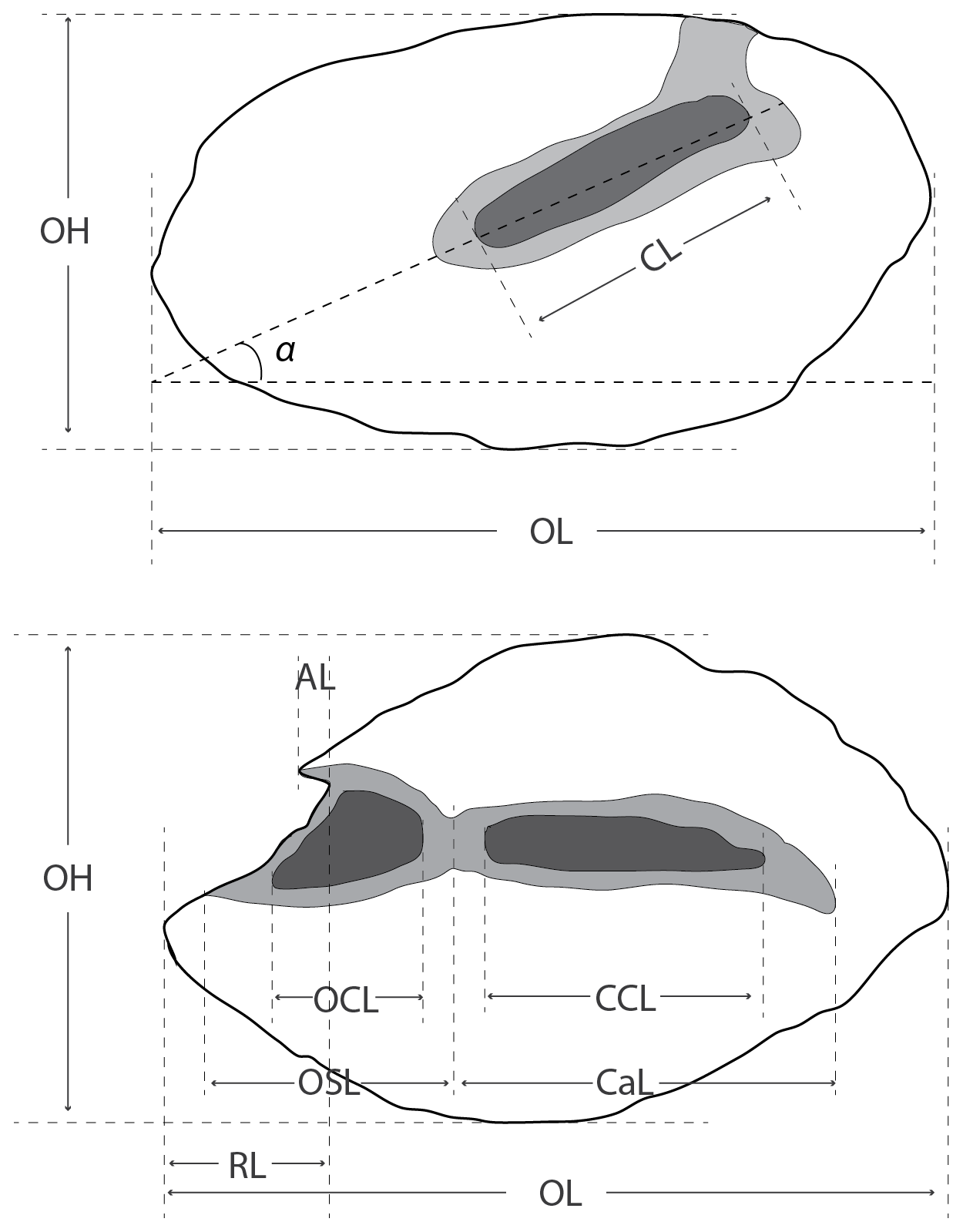
Figure 2Schematic diagram of the measurements obtained on the otoliths. OL, otolith length; OH, otolith height; AL, antirostrum length; RL, rostrum length; CL, colliculum length; OCL, ostial colliculum length; CCL, caudal colliculum length; OSL, ostium length; CaL, caudal length, α, sulcus angle.
In this section, we describe and explain the species identification of the fossil otoliths from the Felli section. The measurements for all identified otoliths are available in the Supplement. The studied material has been deposited at the Athens Museum of Palaeontology and Geology – Vertebrates Collection (AMPG(V)).
-
Infraclass Teleostei Müller, 1845 sensu Arratia, 1999
-
Order Anguilliformes Berg, 1943
-
Family Congridae Kaup, 1856
-
Genus Ariosoma Swainson, 1838
-
Ariosoma mesohellenica sp. nov.
urn:lsid:zoobank.org:act:4B603ED2-93BE-429E-B4EC-8D5F45A9C24C registered on 21/7/2021
Fig. 3A–D
-
2004 Ariosoma balearicum (Delaroche, 1809) – Hoedemakers & Batllori, pl. 1, figs. 8–14.
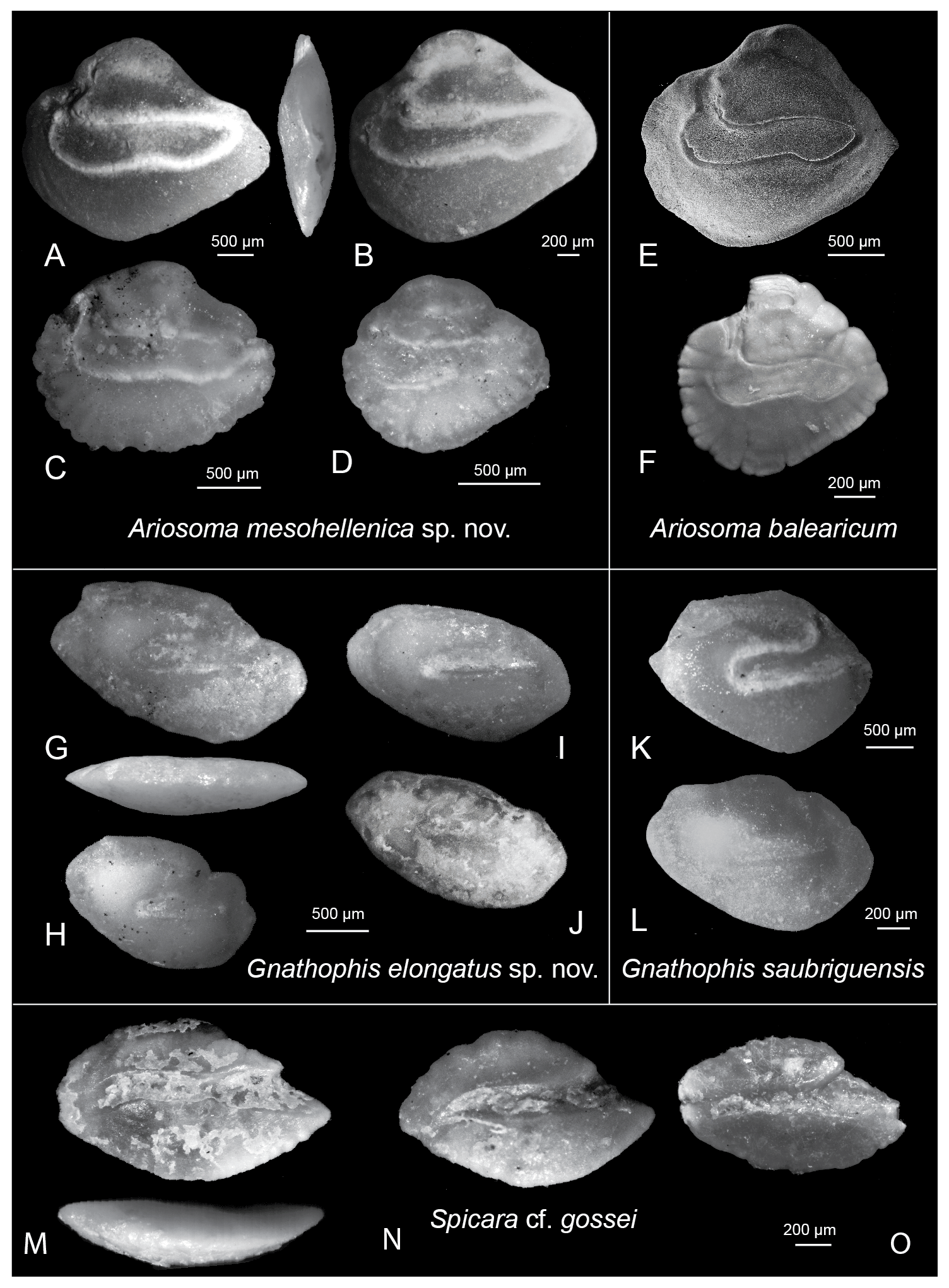
Figure 3Photographs of the identified otoliths from the Felli section. (A–D) Ariosoma mesohellenica sp. nov.: (A) holotype, AMPG(V) 2361, inner face and dorsal view; Felli 11; (B) paratype, AMPG(V) 2364, Felli 12; (C–D) paratypes, 2362 and 2363, Felli 11. (E–F) Ariosoma balearicum (Delaroche), Holocene, Israel. (G–H) Gnathophis elongatus sp. nov., Felli 11: (G) holotype, AMPG(V) 2365, inner face and ventral view; (H–J) paratypes, AMPG(V) 2366–2368. (K–L) Gnathophis saubriguensis (Steurbaut): (K) Felli 11, (L) Felli 12; (M–O) Spicara cf. gossei: (M) inner face and ventral view; Felli 11, (N–O) Felli 12. (C), (H), (K), (L), and (M) have been mirrored to facilitate comparison.
Holotype
Figure 3A, AMPG(V) 2361, Aquitanian, Felli section, level 11, Grevena Prefecture, Greece; OL, 3.40; OH, 2.80; CL, 2.49 mm; α, 4∘.
Paratypes
Two specimens, AMPG(V) 2362–3, same data as holotype; one specimen, AMPG(V) 2364, Aquitanian, Felli section, level 12, Grevena Prefecture, Greece.
Etymology
After the MHB, where the type section sediments were deposited.
Diagnosis
Elongated otoliths with OL/OH of 1.16–1.26. The sulcus extends almost to the distal edge, with CL/OL of 0.58–0.74, and α between 2 and 6∘.
Description
Elongated, thick spindle-shaped otoliths with an elongated central sulcus that opens toward the dorsal rim through an intricate ostial channel. The ostial channel is placed almost at the proximal edge of the sulcus and is perpendicular to it. The dorsal depression is well expressed. The dorsal rim is curved in the distal part.
Distribution
Aquitanian of the western proto-Mediterranean and the MHB.
Remarks
These otoliths have a more elongated sulcus than Ariosoma balearicum otoliths, which almost reaches the posterior edge (Fig. 3E–F; Agiadi and Albano, 2020). Moreover, the dorsal extension is located more frontally, as it is in Ariosoma moravica (Sulc 1932) (Nolf, 2013). The otoliths assigned to A. balearicum from the lower–middle Miocene of Catalonia by Hoedemakers and Batllori (2005) also show a frontal positioning of the dorsal extension. In contrast through to A. moravica, the sulcus is more elongated, indicating that these specimens also should be placed in Ariosoma mesohellenica sp. nov. Furthermore, the specimens assigned to Ariosoma aff. balearicum by Steurbaut (1984) should not be placed within this species because their sulcus is much shorter and does not extend close to the distal rim. Similarly, the otoliths assigned by the same authors to Ariosoma globosum Steurbaut, 1984 are distinguished from those of A. mesohellenica because their sulcus does not extend toward the distal rim and their shape is pointed.
-
Genus Gnathophis Kaup, 1860
-
Gnathophis elongatus sp. nov.
urn:lsid:zoobank.org:act:EF4B2DCC-4246-45AB-840E-1BC29C675410 registered on 21/7/2021
Fig. 3G–J
Holotype
Figure 3G, AMPG(V) 2365, Aquitanian, Felli section, level 11, Grevena Prefecture, Greece; OL, 2.62; OH, 1.44; CL, 1.170; α, 12∘.
Paratypes
Nine specimens, AMPG(V) 2366–74, same data as holotype.
Etymology
Due to the distinct elongated shape of its otoliths.
Diagnosis
Elongated otoliths with OL/OH of 1.60–2.06. The angle of the sulcus relative to the otolith length plane is 10–20∘.
Description
These otoliths are elongated with a long sulcus that forms an angle to the otolith length direction, which opens posterodorsally through a wavy ostial channel. The posterior edge is slightly protruding dorsally. The ventral rim forms an angle.
Distribution
Aquitanian of the proto-Mediterranean Basin.
Remarks
The otoliths of Gnathophis elongatus are elongated and more similar to Conger, unlike Pliocene–Holocene specimens of Gnathophis mystax that are more quadrate (Tuset et al., 2008; Agiadi et al., 2020, 2019, 2018). However, the orientation of the sulcus forms an angle with the otolith length direction, as in Gnathophis species. The otolith figured as Gnathophis catalinensis (Wade) by Steurbaut (1984), now accepted as a synonym of Gnathophis cinctus (Garman) (Grove and Lavenberg, 1997), also displays the same elongate shape, distinct sulcus-to-otolith length angle as those described here. However, the sulcus is much narrower in G. elongatus, making it distinguishable from G. cinctus. Compared to G. saubriguensis (Steurbaut), G. elongatus has a much more elongated shape, the posterodorsal edge is protruding, and the distal–ventral angle is not pointed.
-
Gnathophis saubriguensis (Steurbaut, 1979)
Fig. 3K–L
-
1979 “genus aff. Lemkea” saubriguensis – Steurbaut, pl. 1, figs. 8–12
-
1984 Gnathophis saubriguensis (Steurbaut, 1979) – Steurbaut, pl. 3, figs. 7–8
-
2004 Gnathophis saubriguensis (Steurbaut, 1979) – Nolf and Brzobohaty, pl. 1, fig. 3–5
Material
Fifteen specimens from level 11 and three specimens from level 12.
Distribution
Aquitanian of the Aquitaine Basin (SW France; Steurbaut, 1979, 1984) and the MHB, and Burdigalian of northern Italy (Nolf and Brzobohaty, 2004).
Remarks
The orientation of the sulcus relative to the posterior–anterior axis is angular, with an 11–24∘ angle. The anterior edge shows the same opening feature as the specimens from the late Burdigalian of Piedmont, Italy (Nolf and Brzobohaty, 2004). The distal–ventral angle is pointed, as in the specimens figured from the late Oligocene–lower Miocene of France (Steurbaut, 1984).
-
Order Stomiiformes Regan, 1909
-
Family Phosichthyidae Weitzman, 1974
-
Genus Vinciguerria Jordan & Evermann, 1896
-
Vinciguerria sp.
Material
Two otoliths from level 11 and one otolith from level 12.
Remarks
These otoliths resemble Vinciguerria lucetia (Garman) in the shape and angle of the ventral rim. However, they are too eroded for confident specific identification.
-
Order Myctophiformes Regan, 1911
-
Family Myctophidae Gill, 1893
-
Genus Diaphus Eigenmann & Eigenmann, 1890
-
Diaphus sp.
Material
Five otoliths from level 11.
Remarks
The state of preservation of these otoliths does not permit their identification to species level.
-
Genus Lobianchia Gatti, 1904
-
Lobianchia sp.
Fig. 4A
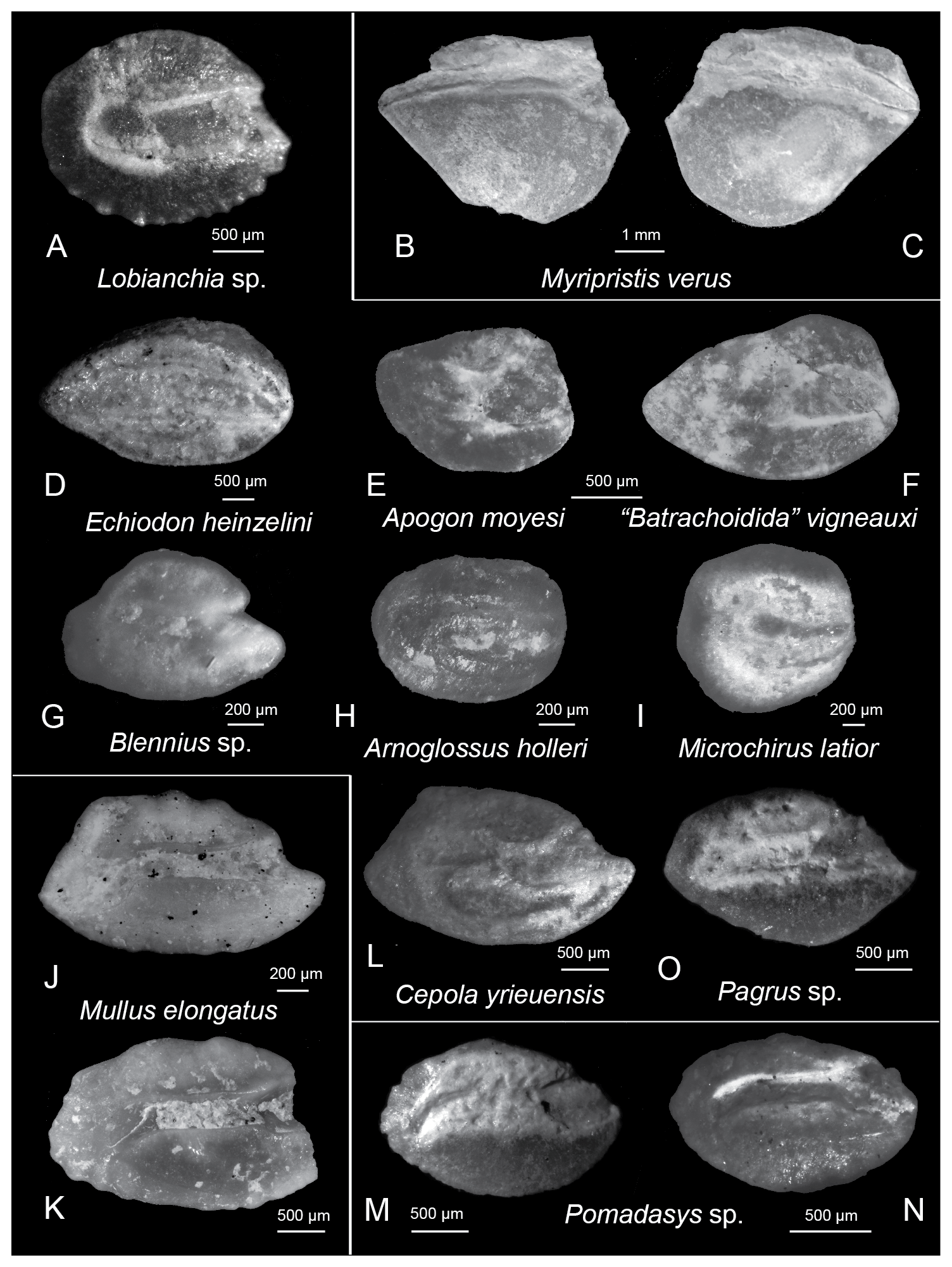
Figure 4Photographs of the identified otoliths from the Felli section. (A) Lobianchia sp., Felli 11; (B–C) Myripristis verus Steurbaut, (B) Felli 11, (C) Felli 12; (D) Echiodon heinzelini Huyghebaert and Nolf, Felli 11; (E) Apogon moyesi Steurbaut, Felli 11; (F) “Batrachoidida” vigneauxi (Steurbaut), Felli 11; (G) Blennius sp., Felli 11; (H) Arnoglossus holleri Weinfurter, Felli 11; (I) Microchirus latior (Schubert), Felli 11; (J–K) Mullus elongatus Steurbaut, (J) Felli 11, (K) Felli 12; (L) Cepola yrieuensis Steurbaut, Felli 11; (M–N) Pomadasys sp., Felli 11; (O) Pagrus sp., Felli 11. (F), (H), (J), and (K) have been mirrored to facilitate comparison.
Material
One otolith from level 11.
Remarks
Compared to Lobianchia dofleinoides Steurbaut, the otolith found here shows neither an anterodorsal curvature of the rim nor a strong dorsal depression. Moreover, the ostium is separated from the cauda with a clearly angular boundary, not one that is perpendicular to the posterodistal direction, as is the case for L. dofleinoides. Nevertheless, the sulcus, especially the ostium part, is quite wide, resembling more L. dofleinoides than L. dofleini (Zugmayer). Thus, it was not possible to assign this specimen to either species, despite similarities, and we therefore left it in open nomenclature.
-
Order Beryciformes Regan, 1909
-
Family Holocentridae Richardson, 1846
-
Genus Myripristis Cuvier, 1829
-
Myripristis verus Steurbaut, 1979
Fig. 4B–C
-
1979 Myripristis verus – Steurbaut, pl. 7, figs. 3–4
-
1984 Myripristis verus Steurbaut, 1979 – Steurbaut, pl. 15, figs. 15–17
Material
One otolith from level 11 and one otolith from level 12.
Distribution
Aquitanian–Burdigalian of SW France (Steurbaut, 1979, 1984) and Aquitanian of the MHB.
Remarks
Myripristis verus otoliths are not as high as those of Myripristis planodorsalis Schwarzhans, 1994. Compared to the “genus Myripristinarum” banaticus Weiler (Steurbaut, 1979) and the “genus Myripristinarum” sp. figured by Steurbaut (1984), the inner face of M. verus is more triangular, and its sulcus is more strongly bending ventrally.
-
Order Ophidiiformes Berg, 1937
-
Family Carapidae Poey, 1867
-
Genus Echiodon Thompson, 1837
-
Echiodon heinzelini Huyghebaert and Nolf, 1979
Fig. 4D
-
1979 Echiodon heinzelini – Huyghebaert and Nolf, p. 74, fig. 5
-
1984 Echiodon heinzelini Huyghebaert and Nolf, 1979 – Steurbaut, pl. 13, figs. 1–6
-
2004 Echiodon heinzelini Huyghebaert and Nolf, 1979 – Nolf and Brzobohaty, pl. 8, fig. 6
Material
One otolith from level 11.
Distribution
Lower Miocene of Belgium (Huyghebaert and Nolf, 1979), SW France (Steurbaut, 1984), northern Italy (Nolf and Brzobohaty, 2004), and the MHB.
Remarks
The sulcus reaches both the proximal and almost the distal tip, similar to E. heinzelini from the lower Miocene of Belgium (Huyghebaert and Nolf, 1979), the Oligocene–lower Miocene of SW France (Steurbaut, 1984), and the Aquitanian of Piedmont, Italy (Nolf and Brzobohaty, 2004), and unlike “genus Neobythitinorum” boulangeri Nolf (Steurbaut, 1984) and the modern species Echiodon dentatus (Nolf, 1980).
-
Order Batrachoidiformes
-
Family Batrachoididae
-
“Batrachoidida” vigneauxi (Steurbaut, 1984)
Fig. 4F
-
1984 “genus Apogoninarum” vigneauxi – Steurbaut, pl. 20, figs. 14–19
-
2013 “Batrachoidida” vigneauxi (Steurbaut, 1984) – Nolf, pl. 144.
Material
Two otoliths from level 11.
Distribution
Aquitanian of SW France (Steurbaut, 1984) and the MHB.
Remarks
These otoliths are elongate with a higher anterior than posterior part and a prominent anterodorsal high. The sulcus is divided into a large, oval ostium that almost reaches the anterior rim, and a smaller, also oval-shaped cauda that is situated slightly dorsally. The specimens examined here resemble very much those figured by Steurbaut (1984) from the Oligocene–lower Miocene of France and reassigned by Nolf (2013) to Batrachoididae.
-
Order Kurtiformes Jordan, 1923
-
Family Apogonidae Günther, 1859
-
Genus Apogon Lacepède, 1801
-
Apogon moyesi Steurbaut, 1982
Fig. 4E
-
1982 Apogon moyesi – Steurbaut, pl. 2, figs. 14–18
-
1984 Apogon moyesi Steurbaut, 1982 – Steurbaut, pl. 20, figs. 20–22
Material
One otolith from level 11.
Distribution
Aquitanian of SW France (Steurbaut, 1982) and the MHB.
Remarks
This specimen is significantly shorter than those assigned to Apogon vigneauxi (Steurbaut, 1984), and it shows the same dorsal depression and relative size of the ostium to the cauda (OSL : CaL = 1.128) as A. moyesi from the Oligocene–Miocene of Aquitaine Basin (Steurbaut, 1982, 1984). Compared to the modern species Apogon imberbis from the Pliocene–Holocene of the eastern Mediterranean Sea (Agiadi et al., 2019, 2020; Agiadi and Albano, 2020), this specimen is shorter (OL : OH = 1.33 compared to a mean of 1.42 in A. imberbis).
-
Order Blenniiformes Bleeker, 1860
-
Family Blenniidae Rafinesque, 1810
-
Genus Blennius Linnaeus, 1758
-
Blennius sp.
Fig. 4G
Material
One otolith from level 11.
Remarks
This otolith is triangular in shape with a sulcus divided into an oval-shaped ostium and a thinner but approximately equal in length oval cauda that slightly turns towards the dorsal side. These characteristics make this specimen similar to “Blennida” bicipitis Steurbaut from the Rupelian of the Aquitaine Basin (Nolf, 2013). However, the specimen examined here has a square posterior rim.
-
Order Pleuronectiformes Bleeker, 1859
-
Family Bothidae Smitt, 1892
-
Genus Arnoglossus Bleeker, 1862
-
Arnoglossus holleri Weinfurter, 1952
Fig. 4H
-
1952 Arnoglossus holleri sp. nov. – Weinfurter, pl. 4, fig. 4
-
1962 Arnoglossus miocenicus sp. nov. – Weiler, fig. 1, 18a–20
-
1979 Arnoglossus miocenicus Weiler, 1962 – Steurbaut, pl. 11, figs. 12–14
-
1980 Arnoglossus sp. – Nolf and Cappetta, pl. 3, figs. 15–16
-
1981 Arnoglossus sp. – Steurbaut & Jonet, pl. 4, fig. 6
-
1984 Arnoglossus sp. – Steurbaut, pl. 34, figs. 17–22
-
1992 Arnoglossus sp. – Radwanska, fig. 158, pl. 38, figs. 1–3
-
1999 Arnoglossus holleri Weinfurter, 1952 – Schwarzhans, p.170–173, figs. 341–352
-
2004 Arnoglossus holleri Weinfurter, 1952 – Nolf and Brzobohaty, pl. 11, fig. 15
Material
One otolith from level 11.
Distribution
Miocene of the Styrian Basin (Weinfurter, 1952), middle Miocene of NW Germany (Weiler, 1962), lower–middle Miocene of the Aquitanian Basin (Steurbaut, 1979, 1984), Miocene of Portugal (Steurbaut and Jonet, 1981), middle Miocene (Badenian) of Poland (Radwanska, 1992), lower Miocene of northern Italy (Nolf and Brzobohaty, 2004) and southern France (Nolf and Cappetta, 1980), and Aquitanian of the MHB.
Remarks
This small otolith is rectangular in shape, with rounded angles. The sulcus is sloping posteroventrally and approximately straight, divided into a long ostium and a rectangular, short cauda. In contrast to the specimen figured by Nolf and Brzobohaty (2004), the otolith found in Felli shows a more regular rectangular shape. Our specimen is quite similar to the modern species Arnoglossus rueppellii (Cocco), but its inner and outer face have the same convexity, whereas A. rueppellii has a flatter inner face (Agiadi and Albano, 2020).
-
Family Soleidae Bonaparte, 1832
-
Genus Microchirus Bonaparte, 1833
-
Microchirus latior (Schubert)
Fig. 4I
-
1906 Solea latior sp. nov. – Schubert, pl. 6, figs. 12–14
-
1979 Solea latior Schubert, 1906 – Anfossi and Mosna, pl. 4, fig. 14
-
1984 Buglossidium frequens – Steurbaut, pl. 35, figs. 9–18
-
1989 Microchirus aff. variegatus (Donovan) – Brzobohaty, pl. 2, fig. 8
-
1989 Buglossidium frequens Steurbaut, 1984 – Brzobohaty, pl. 2, fig. 9
-
1992 Microchirus aff. variegatus (Donovan, 1808) – Radwanska, pl. 38, fig. 9
-
1999 Microchirus latior (Schubert, 1906) – Schwarzhans, p.288, figs. 730–733
-
2004 Buglossidium frequens Steurbaut, 1984 – Nolf & Brzobohaty, pl. 11, figs.13–14
Material
Two otoliths from level 11 and one otolith from level 12.
Distribution
Lower Miocene of Italy (Nolf and Brzobohaty, 2004), the Aquitanian Basin (Steurbaut, 1984), and the MHB. Middle Miocene (Badenian) of Poland (Radwanska, 1992).
Remarks
The otoliths of Microchirus latior are rounded, unlike those of Microchirus variegatus (Donovan, 1808) that have edges on the posterior side (Schwarzhans, 1999; Agiadi et al., 2020).
-
Order Perciformes Bleeker, 1859
-
Family Serranidae Swainson, 1839
-
Genus Mullus Linnaeus, 1758
-
Mullus elongatus Steurbaut, 1984
Fig. 4J–K
-
1984 Mullus elongatus – Steurbaut, pl. 29, figs. 8–12
-
2013 Mullus elongatus Steurbaut, 1984 – Nolf, pl. 285
Material
Two otoliths from level 11 and two otoliths from level 12.
Distribution
Lower Miocene of the Aquitanian Basin (Steurbaut, 1984) and the MHB.
Remarks
These otoliths have a long sulcus, comprised of a small circular ostium and a long cauda, which turns toward the ventral area and opens in the posterior end. The dorsal rim is straight. Compared to the modern species Mullus surmuletus Linnaeus and Mullus barbatus Linnaeus (Hoedemakers and Batllori, 2005; Tuset et al., 2008; Agiadi et al., 2019), the otoliths of M. elognatus are not as high, but more elongated (OL : OH = 1.76–1.82 in M. elongatus but 1.23 in M. barbatus and 1.52 in M. surmulatus, both values from the Pleistocene of the eastern Mediterranean; Agiadi et al., 2019), more so than Mullus bifurcatus Strashimirov reported from the lower–middle Miocene of the Paratethys (Bratishko et al., 2015).
-
Family Cepolidae Rafinesque, 1810
-
Genus Cepola Linnaeus, 1764
-
Cepola yrieuensis Steurbaut, 1984
Fig. 4L
-
1984 Cepola yrieuensis –Steurbaut, pl. 29, figs. 13–17
-
2013 Cepola yrieuensis – Nolf, pl. 294
Material
Two otoliths from level 11.
Distribution
Lower Miocene of the Aquitanian Basin (Steurbaut, 1984) and the MHB. Priabonian–lower Oligocene of SE France (Nolf and Girone, 2008; Girone and Nolf, 2009).
Remarks
These otoliths are boat-shaped and have open, divided sulcus. The ostium is oval–rectangular and has an oval colliculum, whereas the cauda is oval-shaped, with an oval colliculum, and placed more dorsally than the ostium. There is a pronounced ventral fissure. These specimens have distinct posterior and posterodorsal angles that only appear on the otoliths of C. yrieuensis previously reported from the Rupelian of France (Steurbaut, 1984). Compared to C. macrophthalma, these otoliths have an oval, rather than circular cauda, which is placed dorsally. Cepola aff. macrophthalma (Linnaeus) from the middle Miocene of Austria (Nolf, 2013) also has an oval-shaped cauda but lacks a posterodorsal angle. On the contrary, this angle is present in C. robusta Nolf from the Ypresian of France (Nolf, 2013), but only in the young specimens.
-
Order Spariformes Bleeker, 1876
-
Family Sparidae Rafinesque, 1810
-
Genus Pomadasys Rafinesque, 1810
-
Pomadasys sp.
Fig. 4M–N
Material
Six otoliths from level 11.
Remarks
These otoliths are oval-shaped, with a small circular–oval ostium and a long straight cauda. The ventral area is distinctly larger than the dorsal area, and its rim is rounded.
-
Genus Pagellus Valenciennes, 1830
-
Pagellus sp.
Material
One otolith from level 11 and one otolith from level 12.
Remarks
These otoliths are fusiform in shape with a well-curved ventral rim and a sulcus divided into a triangular ostium and a long straight cauda, which is curved at the posterior end.
-
Genus Pagrus Cuvier, 1816
-
Pagrus sp.
Fig. 4O
Material
One otolith from level 11.
Remarks
This otolith is characterized by a pronounced ventral angle. The sulcus is divided into a triangular ostium and a long cauda, which curves at its posterior end. The dorsal area forms a clear angle in the anterior part. There is some resemblance to Pagrus sp. 1 figured by Steurbaut (1984).
-
Genus Spicara Rafinesque, 1810
-
Spicara cf. gossei Steurbaut, 1984
Fig. 3M–O
Material
Nine otoliths from level 11 and seven otoliths from level 12.
Remarks
The general shape of the otoliths is fusiform. They have a straight sulcus and a triangular ostium, which is characteristic of all Spicara spp., but these specimens are particularly angular in shape and have a strongly delineated sulcus. The rostrum is more robust in these specimens than in the modern species Spicara flexuosa, S. smaris, and S. maena (Agiadi and Albano, 2020), and S. gossei from the Oligocene–lower Miocene of France (Steurbaut, 1984). In addition, the posterior end of the cauda is very pointed in the examined specimens.
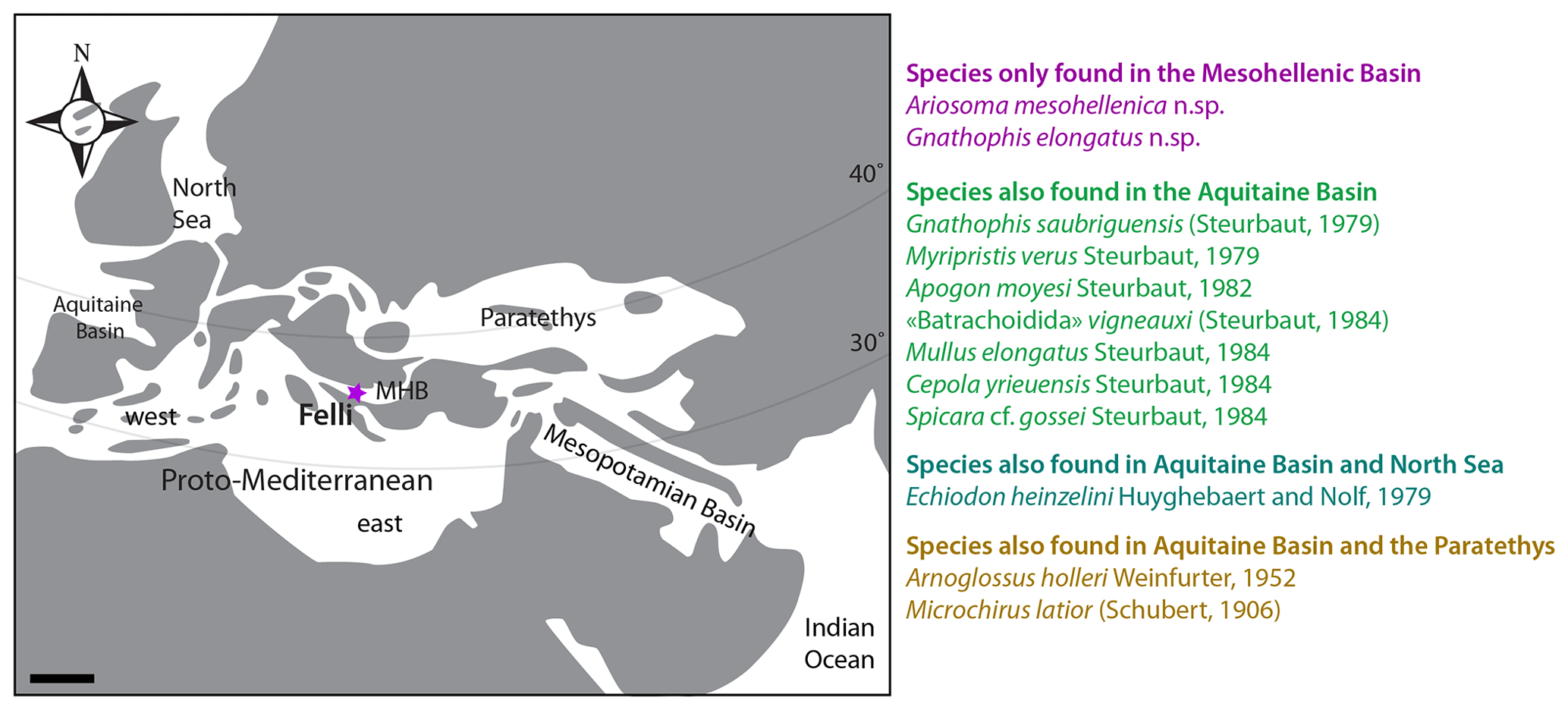
Figure 5Paleogeographic map of the proto-Mediterranean, showing its connections to the Paratethys, Atlantic, and Indo-Pacific realms, and the location of the Mesohellenic Basin (MHB) and the Felli section (modified after early Burdigalian scheme of Popov et al., 2004), and list of the species identified at Felli color-coded to indicate their paleobiogeographic distribution.
5.1 Aquitanian fish biogeography
The fish fauna of the Aquitanian MHB is quite diverse. We identified 19 species from 17 genera, including 2 new species: Ariosoma mesohellenica and Gnathophis elongatus. Overall, the identified fish fauna comprises several species of congrid eels, flatfishes, and sparids, along with lanternfishes, reflecting a shelf environment with easy access to the open ocean. The great diversification of congrids especially reflects a variety of microhabitats along this shelf.
The geological events that led to the formation of the proto-Mediterranean have shaped the distribution of marine taxa (Hou and Li, 2018; Worm and Tittensor, 2018). In the area of the Western Tethys, the late Eocene–middle Miocene MHB (Fig. 5) constituted a marine corridor at the border between the proto-Mediterranean and the Paratethys (Rögl, 1998); the proto-Mediterranean Sea developed due to the closure of the western part of the Tethys Ocean (Bialik et al., 2019; Torfstein and Steinberg, 2020). At this time, the proto-Mediterranean Sea, including the MHB, transited from a global biodiversity hotspot to a subtropical marine sea (Renema et al., 2008). Therefore, any faunal exchanges between the proto-Mediterranean, the Paratethys, the North Sea, the Atlantic, and the Indo-Pacific realms would be expressed in this study area.
Possibly due to the warm Aquitanian climate (Zachos et al., 2001; Westerhold et al., 2020), the distribution ranges of North Sea and NE Atlantic fish species included the MHB in the south. Indeed, this is supported by the distribution of the identified fish species (Fig. 5). Most fish species in the MHB were also present in the NE Atlantic, and, in fact, Echiodon heinzelini was also present in the North Sea. Moreover, the flatfishes Arnoglossus holleri and Microchirus latior were also reported in the Paratethyan realm, which is understandable considering the close proximity between these areas. We do not find any exclusively Indo-Pacific taxa in our fossil assemblage. However, we must note that data on the Miocene fishes of the Indo-Pacific realm are scarce; data outside the Mediterranean and the Atlantic exist mostly from New Zealand (Grenfell, 1984; Schwarzhans et al., 2017). This knowledge gap hampers any further evaluation of the paleobiogeographic distribution of the identified species.
5.2 Comparison with the paleobiogeography of the accompanying molluscan fauna
In the early Neogene, the Mediterranean–Iranian biogeographical province of mollusks (Harzhauser et al., 2002) covered part of the Western Tethys region. The molluscan fauna in this province had a similar taxonomic composition with those of the North Atlantic of France and the Paratethys (Harzhauser et al., 2002; Lozouet, 2014), as in the case of fishes. Shallow marine faunas from the Indo-West Pacific show high degrees of endemism in the early Miocene (Harzhauser et al., 2007, 2017), whereas a turnover in the composition of nearshore mollusks is evident in the Oligocene to early Miocene in the Indian Ocean (Harzhauser et al., 2009).
The molluscan fauna of the Felli section is most similar to shallow marine and coastal assemblages from the NE Atlantic and the Paratethys (Thivaiou et al., 2019). However, in contrast to fishes (for which we do not have an Indo-Pacific fossil record to compare with), the molluscan fauna of the Felli section also includes genera that are characteristic of subtropical to tropical environments (Houbrick, 1991; Janssen et al., 2011), such as the widespread coastal mudflat Terebralia (Thivaiou et al., 2021), currently inhabiting only the Indo-West Pacific, and Finella, which has recently re-entered the Mediterranean Sea from the Red Sea after the opening of the Suez Canal (Albano et al., 2021). This observation might suggest that mollusk and fish distribution ranges may not be similarly affected by paleoenvironmental change, although such a conclusion is in both cases heavily affected by sampling and study bias.
The fish fauna of the early Miocene MHB was in general influenced by the connectivity of the proto-Mediterranean to its surrounding marine basins. The early Miocene fish fauna of the proto-Mediterranean, as revealed by the otolith assemblage of the Felli section, was a mixture of Atlantic, Paratethyan, North Sea, and Mediterranean endemic elements. Unfortunately, the fossil record so far for this interval has been scarce. Nevertheless, the present findings offer some insight into the early Miocene evolution of the Mediterranean marine fish fauna. Future research should aim to fill in the considerable gaps in the record for the early–middle Miocene, especially in the Indo-Pacific realm.
The supplementary data are available open-access with this article.
The supplement related to this article is available online at: https://doi.org/10.5194/fr-24-233-2021-supplement.
KA was responsible for data curation, formal analysis, investigation, methodology, resources, and writing; DT for conceptualization, funding acquisition, investigation, methodology, resources, and writing; and EK for investigation, resources, and writing.
The authors declare that they have no conflict of interest.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This research has been co-financed by Greece and the European Union (European Social Fund-ESF) through the operational program “Human Resources Development, Education and Lifelong Learning 2014–2020” in the context of the project “Mollusc and fish migrations in the dynamic environments of the early to middle Miocene in the Mediterranean” (MIS 5047960). The authors would like to thank Gary Stringer and the anonymous reviewer for their constructive comments.
This research has been supported by the Ministry of National Education and Religious Affairs (grant no. MIS 5047960).
This paper was edited by Florian Witzmann and reviewed by Gary Stringer and one anonymous referee.
Agiadi, K. and Albano, P. G.: Holocene fish assemblages provide baseline data for the rapidly changing eastern Mediterranean, Holocene, 30, 1438–1450, https://doi.org/10.1177/0959683620932969, 2020.
Agiadi, K., Triantaphyllou, M., Girone, A., and Karakitsios, V.: The early Quaternary palaeobiogeography of the eastern Ionian deep-sea Teleost fauna: A novel palaeocirculation approach, Palaeogeogr. Palaeocl., 306, 228–242, https://doi.org/10.1016/j.palaeo.2011.04.029, 2011.
Agiadi, K., Antonarakou, A., Kontakiotis, G., Kafousia, N., Moissette, P., Cornée, J.-J., Manoutsoglou, E., and Karakitsios, V.: Connectivity controls on the late Miocene eastern Mediterranean fish fauna, Int. J. Earth Sci., 106, 1147–1159, https://doi.org/10.1007/s00531-016-1355-7, 2017.
Agiadi, K., Girone, A., Koskeridou, E., Moissette, P., Cornée, J.-J., and Quillévéré, F.: Pleistocene marine fish invasions and paleoenvironmental reconstructions in the eastern Mediterranean, Quaternary Sci. Rev., 196, 80–99, https://doi.org/10.1016/j.quascirev.2018.07.037, 2018.
Agiadi, K., Vasileiou, G., Koskeridou, E., Moissette, P., and Cornée, J.-J.: Coastal fish otoliths from the early Pleistocene of Rhodes (eastern Mediterranean), Geobios, 55, 1–15, https://doi.org/10.1016/j.geobios.2019.06.006, 2019.
Agiadi, K., Giamali, C., Girone, A., Moissette, P., Koskeridou, E., and Karakitsios, V.: The Zanclean marine fish fauna and palaeoenvironmental reconstruction of a coastal marine setting in the eastern Mediterranean, Palaeobiodiversity Palaeoenvironments, 100, 773–792, https://doi.org/10.1007/s12549-019-00404-4, 2020.
Albano, P. G., Steger, J., Bošnjak, M., Dunne, B., Guifarro, Z., Turapova, E., Hua, Q., Kaufman, D. S., Rilov, G., and Zuschin, M.: Native biodiversity collapse in the eastern Mediterranean, P. Roy. Soc. B Biol. Sci., 288, 20202469, https://doi.org/10.1098/rspb.2020.2469, 2021.
Bialik, O. M., Frank, M., Betzler, C., Zammit, R., and Waldmann, N. D.: Two-step closure of the Miocene Indian Ocean Gateway to the Mediterranean, Sci. Rep.-UK, 9, 1–10, https://doi.org/10.1038/s41598-019-45308-7, 2019.
Bratishko, A., Schwarzhans, W., Reichenbacher, B., Vernyhorova, Y., and Ćorić, S.: Fish otoliths from the Konkian (Miocene, early Serravallian) of Mangyshlak (Kazakhstan): testimony to an early endemic evolution in the Eastern Paratethys, Palaontol. Z., 89, 839–889, https://doi.org/10.1007/s12542-015-0274-4, 2015.
Brunn, J. H.: Contribution a l'etude geologique du Pinde septentrional et d'une partie de la Macedonie occidentale, Ann. Geol. Pays Hell., 7, 1–358, 1956.
Cornacchia, I., Agostini, S., and Brandano, M.: Miocene Oceanographic Evolution Based on the Sr and Nd Isotope Record of the Central Mediterranean, Paleoceanogr. Paleocl., 33, 31–47, https://doi.org/10.1002/2017PA003198, 2018.
Fermeli, G. and Ioakim, C.: Biostratigraphy and palaeoecological interpretation of Miocene successions in the molassic deposits of Tsotylion, Mesohellenic Trench (Grevena area, northern Greece), Paleontol. Evolucio, 24–25, 199–208, 1992.
Ferrière, J., Reynaud, J.-Y., Pavlopoulos, A., Bonneau, M., Migiros, G., Chanier, F., Proust, J.-N., and Gardin, S.: Geologic evolution and geodynamic controls of the Tertiary intramontane piggyback Meso-Hellenic basin, Greece, Bull. Société Géologique Fr., 175, 361–381, https://doi.org/10.2113/175.4.361, 2004.
Ferrière, J., Chanier, F., Reynaud, J., Pavlopoulos, A., Ditbanjong, P., and Coutand, I.: Evolution of the Mesohellenic Basin (Greece): a synthesis, J. Virtual Explor., 45, 1–51, 2013.
Girone, A. and Nolf, D.: Fish otoliths from the Priabonian (Late Eocene) of North Italy and South-East France – Their paleobiogeographical significance, Rev. Micropaleontol., 52, 195–218, https://doi.org/10.1016/j.revmic.2007.10.006, 2009.
Grenfell, H. R.: Early Miocene teleost otoliths from Parengarenga Harbour, New Zealand, N.Z. J. Geol. Geophys., 27, 51–96, https://doi.org/10.1080/00288306.1984.10422291, 1984.
Grove, J. S. and Lavenberg, R. J.: The fishes of the Galapagos Islands. Stanford University Press, Standford, 863 pp., 1997.
Harzhauser, M., Piller, W. E., and Steininger, F. F.: Circum-Mediterranean Oligo–Miocene biogeographic evolution – the gastropods' point of view, Palaeogeogr. Palaeocl., 183, 103–133, https://doi.org/10.1016/S0031-0182(01)00464-3, 2002.
Harzhauser, M., Kroh, A., Mandic, O., Piller, W. E., Göhlich, U., Reuter, M., and Berning, B.: Biogeographic responses to geodynamics: A key study all around the Oligo-Miocene Tethyan Seaway, Zool. Anz., 246, 241–256, https://doi.org/10.1016/j.jcz.2007.05.001, 2007.
Harzhauser, M., Reuter, M., Piller, W. E., Berning, B., Kroh, A., and Mandic, O.: Oligocene and Early Miocene gastropods from Kutch (NW India) document an early biogeographic switch from Western Tethys to Indo-Pacific, Palaeontol. Z., 83, 333–372, https://doi.org/10.1007/s12542-009-0025-5, 2009.
Harzhauser, M., Reuter, M., Mohtat, T., and Piller, W. E.: Early Miocene reef- and mudflat-associated gastropods from Makran (SE-Iran), PalZ, 91, 519–539, https://doi.org/10.1007/s12542-017-0354-8, 2017.
Hoedemakers, K. and Batllori, J.: Fish otoliths from the early and middle Miocene of the Penedès (Catalunya, Spain), Batalleria, 12, 105–134, 2005.
Hou, Z. and Li, S.: Tethyan changes shaped aquatic diversification, Biol. Rev., 93, 874–896, https://doi.org/10.1111/brv.12376, 2018.
Houbrick, R. S.: Systematic review and functional morphology of the mangrove snails Terebralia and Telescopium (Potamididae; Prosobranchia), Malacologia, 33, 1–2, 289–338, 1991.
Huyghebaert, B. and Nolf, D.: Otolithes de Teleosteens et biostratigraphie des Sables de Zonderschot (Miocene Moyen de la Belgique), Meded. Van Werkgr. Voor Tert. En Kwartaire Geol., 16, 59–100, 1979.
Janssen, R., Zuschin, M., and Baal, C.: Gastropods and their habitats from the northern Red Sea (Egypt: Safaga). Part 2: Caenogastropoda: Sorbeoconcha and Littorinimorpha, Ann. Naturhistorischen Mus. Wien Ser. A, 113, 373–509, 2011.
Kilias, A., Vamvaka, A., Falalakis, G., Sfeikos, A., Papadimitriou, E., Gkarlaouni, C., and Karakostas, V.: The Mesohellenic trough and the Paleogeone Thrace Basin on the Rhodope Massif, their structural evolution and geotectonic significance in the Hellenides, J. Geol. Geophys., 4, 198, https://doi.org/10.4172/2329-6755.1000198, 2015.
Lozouet, P.: Temporal and latitudinal trends in the biodiversity of European Atlantic Cenozoic gastropod (Mollusca) faunas. A base for the history of biogeographic provinces, Carnets Geol.-Noteb. Geol., 2014.
Mavridis, A., Kelepertzis, A., Tsaila-Monopolis, S., Skourtsi-Koroneou, V., and Moores, H.: Geological map of Greece scale, Knidi sheet, Institute of Geology and Mineral Exploration, Athens, 1985.
Nelson, J. S., Grande, T. C., and Wilson, M. V. H.: Fishes of the World, 5th Edn., Wiley, 752 pp., 2016.
Nolf, D.: Etude monographique des otolithes des Ophidiiformes actuels et revision des especes fossiles (Pisces, Teleostei), Meded. Van Werkgr. Voor Tert. En Kwartaire Geol., 17, 71–195, 1980.
Nolf, D.: The diversity of fish otoliths, past and present, Royal Belgian Institute of Natural Sciences, Brussels, 222 pp., pl. 359, 2013.
Nolf, D. and Brzobohaty, R.: Otolithes de poissons du Miocene Inferieur Piemontais, Riv. Piemontese Storia Nat., 25, 69–118, 2004.
Nolf, D. and Cappetta, H.: Les otolithes de Teleosteens du Miocene de Montpeyroux (Herault, France), Palaeovertebrata, 10, 1–28, 1980.
Nolf, D. and Girone, A.: Early Oligocene fish otoliths from the Castellane area (SE France) and an overview of Mediterranean teleost faunas at the Eocene-Oligocene boundary, Neues Jahrb. Geol. P.-A., 248, 139–157, https://doi.org/10.1127/0077-7749/2008/0248-0139, 2008.
Popov, S. V., Rögl, F., Rozanov, A. Y., Steininger, F. F., Shcherba, I. G., and Kovac, M.: Lithological-paleogeographic maps of Paratethys, Cour Forsch – Inst Senckenberg, 250, 46 pp., 2004.
Radwanska, U.: Fish otoliths in the Middle Miocene (Badenian) deposits of southern Poland, Acta Geol. Pol., 42, 141–328, 1992.
Reichenbacher, B.: A partly endemic euryhaline fish fauna (otoliths, teeth) from the Early Miocene of the Aix-Basin (Provence, southern France), CFS Cour. Forschungsinstitut Senckenberg, 113–127, 2004.
Reichenbacher, B. and Cappetta, H.: First evidence of an early Miocene marine teleostean fish fauna (otoliths) from La Paillade (Montpellier, France), Palaeovertebrata, 28, 1–46, 1999.
Renema, W., Bellwood, D. R., Braga, J. C., Bromfield, K., Hall, R., Johnson, K. G., Lunt, P., Meyer, C. P., McMonagle, L. B., Morley, R. J., O'Dea, A., Todd, J. A., Wesselingh, F. P., Wilson, M. E. J., and Pandolfi, J. M.: Hopping Hotspots: Global Shifts in Marine Biodiversity, Science, 321, 654–657, https://doi.org/10.1126/science.1155674, 2008.
Rögl, F.: Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene), Ann. Naturhistorischen Mus. Wien, 99, 279–310, 1998.
Schwarzhans, W.: A comparative morphological treatise of recent and fossil otoliths of the order Pleuronectiformes, Verlag Dr.Friedrich Pfeil, Munchen, 391 pp., 1999.
Schwarzhans, W.: A comparative morphological study of recent otoliths of the Congridae, Muraenesocidae, Nettastomatidae and Colocongridae (Anguilliformes), Memoire Della Soc. Ital. Sci. Nat. E Mus. Storia Nat. Milano, 46, 329–354, 2019.
Schwarzhans, W., Lee, D. E., and Gard, H. J. L.: Otoliths reveal diverse fish communities in Late Oligocene estuarine to deep-water paleoenvironments in southern Zealandia, N. Z. J. Geol. Geophys., 60, 433–464, https://doi.org/10.1080/00288306.2017.1365734, 2017.
Steurbaut, E.: Les otolithes de Téléostéens des Marnes de Saubrigues (Miocène d'Aquitane méridionale, France), Palaeontogr. Beitr. Zur Naturgeschichte Vorz. Paläozoology Stratigr., 1979.
Steurbaut, E.: Les otolithes de teleosteens du gisement de Peyrere a Peyrehorade (couches de passage de l'Oligocene au Miocene d'Aquitaine meridionale, France), Meded. – Werkgr. Voor Tert. En Kwartaire Geol., 19, 37–57, 1982.
Steurbaut, E.: Les otolithes de Téléostéens de l'Oligo-Miocène d'Aquitaine (Sud-Ouest de la France), Palaeontogr. Abt. A, 186, 1–162, 1984.
Steurbaut, E. and Jonet, S.: Revision des otolithes de Teleosteens du Miocene Portugais, Bull. Soc. Belge Geol., 90, 191–229, 1981.
Sun, J., Sheykh, M., Ahmadi, N., Cao, M., Zhang, Z., Tian, S., Sha, J., Jian, Z., Windley, B. F., and Talebian, M.: Permanent closure of the Tethyan Seaway in the northwestern Iranian Plateau driven by cyclic sea-level fluctuations in the late Middle Miocene, Palaeogeogr. Palaeocl., 564, 110172, https://doi.org/10.1016/j.palaeo.2020.110172, 2021.
Thivaiou, D., Harzhauser, M., and Koskeridou, E.: Early Miocene gastropods from the Felli section (Proto-Mediterranean Sea, NW Greece), Geodiversitas, 41, 323–366, 2019.
Thivaiou, D., Harzhauser, M., and Koskeridou, E.: Cenozoic diversity and distribution of the mangrove- and mudflat-associated gastropod genus Terebralia (Potamididae: Caenogastropoda) in Europe, C.R. Palevol., in review, 2021.
Torfstein, A. and Steinberg, J.: The Oligo–Miocene closure of the Tethys Ocean and evolution of the proto-Mediterranean Sea, Sci. Rep.-UK, 10, 13817, https://doi.org/10.1038/s41598-020-70652-4, 2020.
Tuset, V. M., Lombarte, A., and Assis, C. A.: Otolith atlas for the western Mediterranean, north and central eastern Atlantic, Sci. Mar., 72, 7–198, https://doi.org/10.3989/scimar.2008.72s17, 2008.
Weiler, W.: Fisch-Otolithen aus dem oberen Mittelmiozän von Twistringen. Bez. Bremen (NW Deutschland), Geol. Jahrb., 80, 277–294, 1962.
Weinfurter, E.: Die Otolithen der Wetzelsdorfer Schichten und des Florianer Tegels (Miozän, Steiermark), Sitzber Österr Akad Wiss Math-Natw Kl, 161, 455–498, 1952.
Westerhold, T., Marwan, N., Drury, A. J., Liebrand, D., Agnini, C., Anagnostou, E., Barnet, J. S. K., Bohaty, S. M., Vleeschouwer, D. D., Florindo, F., Frederichs, T., Hodell, D. A., Holbourn, A. E., Kroon, D., Lauretano, V., Littler, K., Lourens, L. J., Lyle, M., Pälike, H., Röhl, U., Tian, J., Wilkens, R. H., Wilson, P. A., and Zachos, J. C.: An astronomically dated record of Earth's climate and its predictability over the last 66 million years, Science, 369, 1383–1387, https://doi.org/10.1126/science.aba6853, 2020.
Wielandt-Schuster, U., Schuster, F., Harzhauser, M., Mandic, O., Kroh, A., Rogl, F., Reisinger, J., Liebetrau, V., Steininger, F. F., and Piller, W. E.: Stratigraphy and palaeoecology of Oligocene and Early Miocene sedimentary sequences of the Mesohellenic Basin (NW Greece), Cour. Forsch.-Inst. Senckenberg, 248, 1–55, 2004.
Worm, B. and Tittensor, D. P.: A Theory of Global Biodiversity, Princeton University Press, New Jersey, 214 pp., 2018.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K.: Trends, Rhythms, and Aberrations in Global Climate 65 Ma to Present, Science, 292, 686–693, https://doi.org/10.1126/science.1059412, 2001.
Zelilidis, A., Piper, D. J. W., and Kontopoulos, N.: Sedimentation and basin evolution of the Oligocene-Miocene Mesohellenic basin, Greece, AAPG Bull., 86, 161–182, 2002.