the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 02 Sep 2021
| 02 Sep 2021
A new odontocete (Inioidea, Odontoceti) from the late Neogene of North Carolina, USA
Stephen J. Godfrey
Carolina S. Gutstein
Donald J. Morgan III
A new monotypic genus of Neogene odontocete (Isoninia borealis) is named on the basis of a partial skull (CMM-V-4061). The holotype was found on the riverbed of the Meherrin River (North Carolina, USA) and probably originated from the Miocene marine Eastover Formation. Deep interdigitation of the cranial sutures indicates that this individual was mature. The new taxon differs from all other delphinidans in the unique combination of the following characters: anteriorly retracted premaxillae and maxillae; premaxillae not contacting nasals; thick nasals with ventrolateral margins deeply imbedded within corresponding troughs in the frontals (this is an apomorphy); nasals with transversely convex dorsal surface; nearly symmetrical vertex; os suturarum (or interparietal or extra folds of the frontals) at the vertex; large dorsal infraorbital foramen level with the posterior margin of the external bony nares; and a postorbital recess on the ventrolateral face of the frontal below and behind the postorbital process of the frontal. This odontocete exhibits two small but pronounced concavities on the cerebral face of the frontal/presphenoid that are presumed to have held vestigial olfactory bulbs in life. Inioidea is only diagnosed by a single unequivocal synapomorphy: width across nasals and nares subequal. Isoninia shares this feature with other inioids and forms the basis for the placement of Isoninia within this clade. A relatively high vertex coupled with a supraoccipital that is deeply wedged between the frontals suggests placement of this new inioid species within the family Iniidae. This is the second inioid described from the Eastover Formation (the other being Meherrinia isoni). This new species adds new cranial morphology and a new combination of cranial characters to this taxonomically small but growing group of mostly marine and mostly Western Hemisphere odontocetes (urn:lsid:zoobank.org:pub:E8B817CA-B250-42B3-9365-36EFBFE351C9).
- Article
(6327 KB) -
Supplement
(1263 KB) - BibTeX
- EndNote





The extant South American river dolphins (Inia and Pontoporia) are traditionally grouped in the Inioidea (sensu de Muizon, 1988), a conclusion with which more recent work is in agreement; however, there has been some discussion of whether its sister group is the Yangtze river dolphin (Lipotidae) or modern marine dolphins (Delphinoidea) (Arnason and Guldberg, 1996; Yang and Zhou, 1999; Yang et al., 2005; Cassens et al., 2000; Hamilton et al., 2001; Nikaido et al., 2001; Cozzuol, 2010; Geisler and Sanders, 2003; McGowen et al., 2009; Geisler et al., 2011, 2012; Gatesy et al., 2013; Pyenson et al., 2015; Post et al., 2017; Lambert et al., 2017, 2018, 2020; McGowen et al., 2020). Extant Inioidea are comprised of two monotypic families (Ruiz-Garcia and Shostell, 2010), Iniidae Gray, 1846, and Pontoporiidae Gill, 1871. Inia is a strictly freshwater form that includes three living species: Inia geoffrensis (de Blainville, 1817) found within the Orinoco and Amazon river basins, Inia boliviensis (Banguera-Hinestroza et al., 2002) that inhabits the Beni–Mamoré river basin, and possibly Inia araguaiaensis (Hrbek et al., 2014) from the Araguaia–Tocantins river basin. Pontoporia is monotypic (P. blainvillei; Gervais and d'Orbigny, 1844) and occupies shallow marine and estuarine environments along the southeastern coast of South America, with some entrance into proximal riverine systems including Babitonga Bay, southern Brazil (Cremer and Simões-Lopes, 2005), and the La Plata river of Argentina and Uruguay (Casinos) (Secchi, 2010).
The Pontoporiidae (sensu Geisler et al., 2012) is known from the late middle Miocene to the Holocene. Several authors (Lambert and Post, 2005; Godfrey and Barnes, 2008; Gutstein et al., 2009; Lambert and Muizon, 2013; Pyenson and Hoch, 2007; Pyenson et al., 2015; Post et al., 2017; Lambert et al., 2020) have detailed the geographic and stratigraphic extent of the extinct members of this family, which includes small-bodied, mostly coastal odontocetes with rostra ranging from short to long.
Extinct iniids include Kwanzacetus khoisani Lambert et al., 2018, from the Kwanza Basin, Angola; the Argentinian forms Ischyrorhynchus vanbenedeni Ameghino, 1891 (Cozzuol, 1985, 1996, 2010; Cione et al., 2011; Pyenson et al., 2015), Saurocetes argentinus Burmeister, 1871, and S. gigas Cozzuol, 1989, all from the late Miocene Ituzaingó Formation (Cozzuol, 1989, 1996); a Pleistocene species of Inia from Brazil (Cozzuol, 2010); Isthminia panamensis Pyenson et al., 2015, from the late Miocene Chagres Formation of Panama; Brujadelphis ankylorostris Lambert et al., 2017, from the late Miocene Pisco Formation, Pisco Basin, in Peru; and Goniodelphis hudsoni Allen, 1941, from the late Miocene Bone Valley Formation of Florida, USA (Morgan, 1994).
Within Inioidea, several taxa move from one family to the other in successive phylogenetic analyses, which indicates that for some species final family attribution is not yet settled. Furthermore, eventually some of those could even fall as stem inioids, branching before the node Iniidae + Pontoporiidae.
The discovery of marine iniids has fostered a discussion about the Mio-Pliocene global distribution of inioids in general and has raised questions about the timing and paleobiology of the origin and diversification of marine forms and of the extinction of most iniids except for the exclusively freshwater South American riverine forms (Cassens et al., 2000; Gutstein et al., 2014; Pyenson et al., 2015).
Odontocetes, including inioids, present a dizzying array of cranial morphologies. Our goal here is to add to that diversity by describing and naming a new genus and species of inioid from the late Neogene of North Carolina (USA).
2.1 CT scanning
The holotype partial skull, CMM-V-4061, was CT scanned in the Department of Anthropology at the National Museum of Natural History, The Smithsonian Institution, Washington, D.C., USA. The 3 mm helical CT scans in the transverse plane were taken at 120 kV and 280 mA.
2.2 Whitening and photography
In its natural state, the holotype skull (CMM-V-4061) is very dark, so to improve contrast in Figs. 2 and 3, the specimen was lightly dusted with sublimed ammonium chloride (a whitening technique described by Cooper, 1935, and Feldman, 1989). After the specimen was photographed with a Nikon Coolpix 510 camera on a black velvet background under fluorescent light, the ammonium chloride was removed by holding the specimen under running water (Shelburne and Thompson, 2016).
2.3 Institutional abbreviations
CMM-V, Calvert Marine Museum fossil vertebrate collection, Solomons, Maryland, USA.
MGUH VP, Geological Museum of Copenhagen University, Copenhagen, Denmark.
NMNH, National Museum of Natural History, The Smithsonian Institution, Washington, D.C., USA.
2.4 Anatomical terminology
For the anatomy of the skull, we follow the terminology proposed by Mead and Fordyce (2009) for the extant delphinid Tursiops.
-
CETACEA Brisson, 1762
-
ODONTOCETI Flower, 1867
-
DELPHINIDA de Muizon, 1984
-
Superfamily INIOIDEA de Muizon, 1988
-
Family ?INIIDAE
-
Genus Isoninia gen. nov.
Figs. 2 and 3
Type species
Isoninia borealis, sp. nov., herein designated.
Etymology
The genus name is a combination of “Ison”, the family (surname) name of Ronald Ison, who found and donated the type specimen to the Calvert Marine Museum, and Inia, the feminine generic epithet of the Amazon river dolphin (Inia geoffrensis), also known as the boto or pink river dolphin. The species name “borealis” is Latin for “northern”, a reference to the type and only known specimen having been found in the Northern Hemisphere.
Holotype
The holotype is CMM-V-4061 (Figs. 2 and 3). It consists of an incomplete skull, preserving the posterior facial region, from the lateral margins of the bony nares to the supraoccipital, including the vertex. Ventrally, part of the vomer/presphenoid complex is also preserved. This specimen was alluded to by Geisler et al. (2012) prior to it having been assigned a CMM catalogue number: “Other odontocete cranial elements are known from the same locality. However, they cannot be assigned to the present taxon either because they do not overlap with the aforementioned specimens that constitute the hypodigm or because of the presence of at least one other comparably sized but currently undescribed inioid from the same locality” (Geisler et al., 2012, p. 201).
Type locality and horizon
The holotype was recovered by Ronald Ison (from the same location where the type and referred specimens of Meherrinia isoni were collected; Geisler et al., 2012) from the bed of the Meherrin River – a tributary of the Chowan River, Murfreesboro, Hertford County, North Carolina, USA (near 36∘27′ N, 77∘5′ W). More detailed locality information is available from the authors to qualified individuals upon request. Unfortunately, the specimen was not found in situ and was devoid of entombing sediment, as were all specimens of Meherrinia (Geisler et al., 2012). Nevertheless, it is considered to have been locally derived from the Cobham Bay Member of the Eastover Formation (upper Miocene, Messinian) (Lauck W. Ward, personal communication, 30 November 2009) (Fig. 1). Because it was not collected in situ, it is possible that it could have come from the Pliocene Yorktown Formation, but according to Ward (personal communication, 30 November 2009) this derivation is less likely because of where along the river bed the specimen was recovered. Conservatively, the specimen can be considered to have come from a time interval encompassing the upper Tortonian (Miocene, i.e., the Claremont Manor Member of the Eastover Formation) to the lower Piacenzian (Pliocene). As in Geisler et al. (2012), another argument in favor of the derivation of Isoninia from the Eastover Formation is the observation that specimens of neither Isoninia borealis nor Meherrinia isoni have been found in the Yorktown Formation despite extensive collection and study of the fauna of the Yorktown Formation at the Nutrien Aurora Phosphate Mine (formerly known as the Lee Creek Mine, Aurora, North Carolina) for more than 40 years (Whitmore and Kaltenbach, 2008).
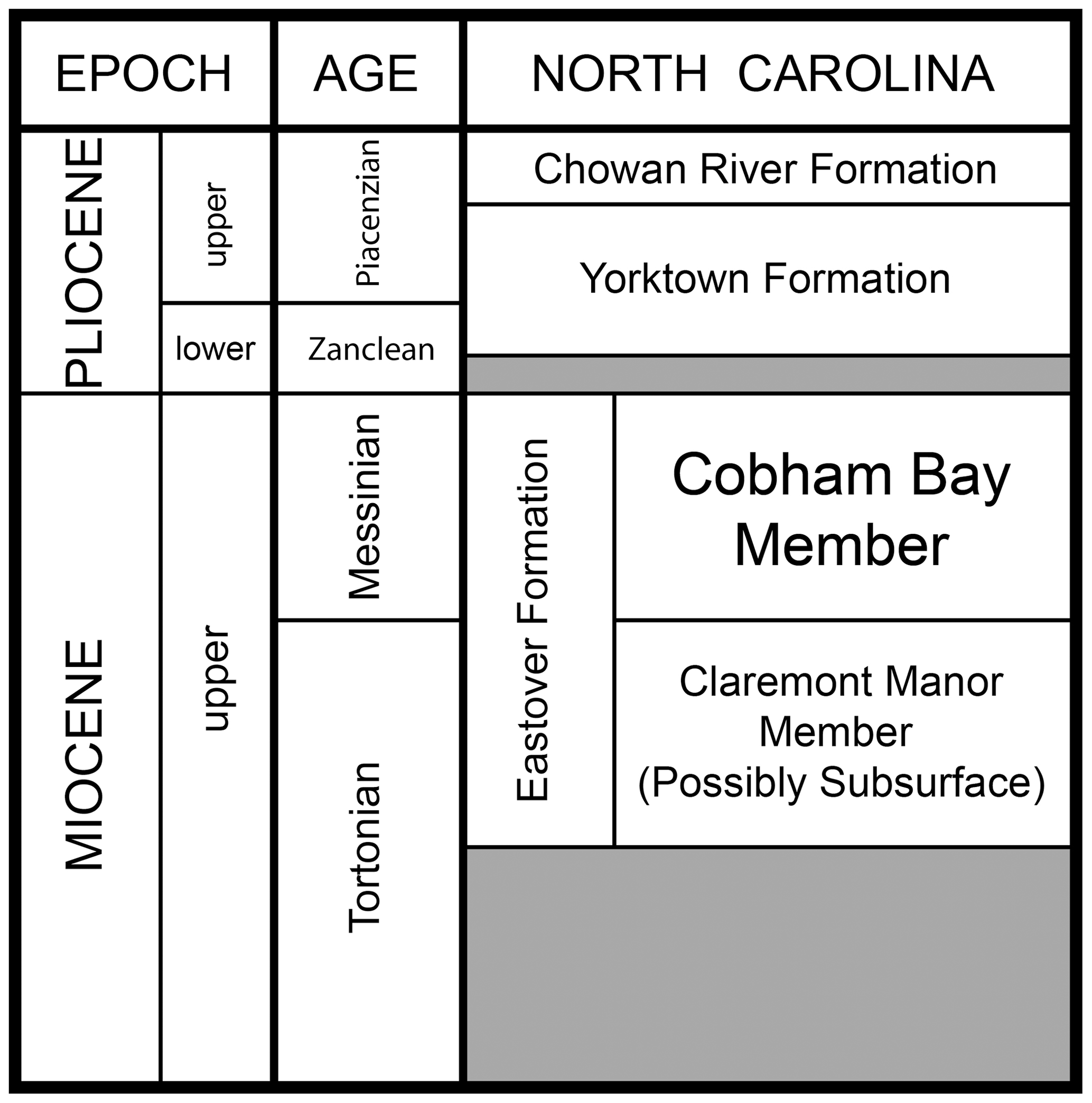
Figure 1Stratigraphy of the marine late Miocene to Pliocene of North Carolina, USA, showing the Cobham Bay Member of the Eastover Formation, from which CMM-V-4061, the holotype and only known specimen of Isoninia borealis (gen. nov., sp. nov.), is thought to be derived. From Geisler et al. (2012), which they modified from Ward and Andrews (2008).
Diagnosis
Isoninia is diagnosed on the basis of the following unique combination of features, most of which are not apomorphic: anteriorly retracted premaxillae and maxillae (i.e., no contact between premaxillae and nasals and maxillae retracted anteriorly from the sides of the frontals below the vertex); large posterior-most dorsal infraorbital foramen at approximately the same level with posterior margin of external bony nares; robust nasals, anteroposteriorly longer than transversely wide in dorsal view with ventrolateral margins turned ventromedially to form ridges deeply imbedded within corresponding troughs in frontals (italicized text represents an apomorphy); nasals large; nasals with transversely convex dorsal surface; dorsal surface of nasals not inflated; elevated, bilaterally compressed nearly symmetrical vertex, narrower than external bony nares; frontals form apex of the vertex; presence of os suturarum (or extra folds of the frontals or a projection of the interparietal) at the vertex; in ventral view, pronounced fossa on ventrolateral face of frontal below and behind postorbital process of frontal for the postorbital lobe of the pterygoid sinus (the postorbital recess).
Description
The bones that are preserved are thickened by comparison to a comparably sized Inia. These thickened bones along with the degree to which the ventral surfaces of the maxillae and nasals deeply suture to the frontals (Fig. 3c) indicate that the holotype of Isoninia borealis was a mature individual (as is also seen in the holotype of the inioid Stenasodelphis russellae, Godfrey and Barnes, 2008, and in the holotype of the iniid Kwanzacetus khoisani, Lambert et al., 2018).

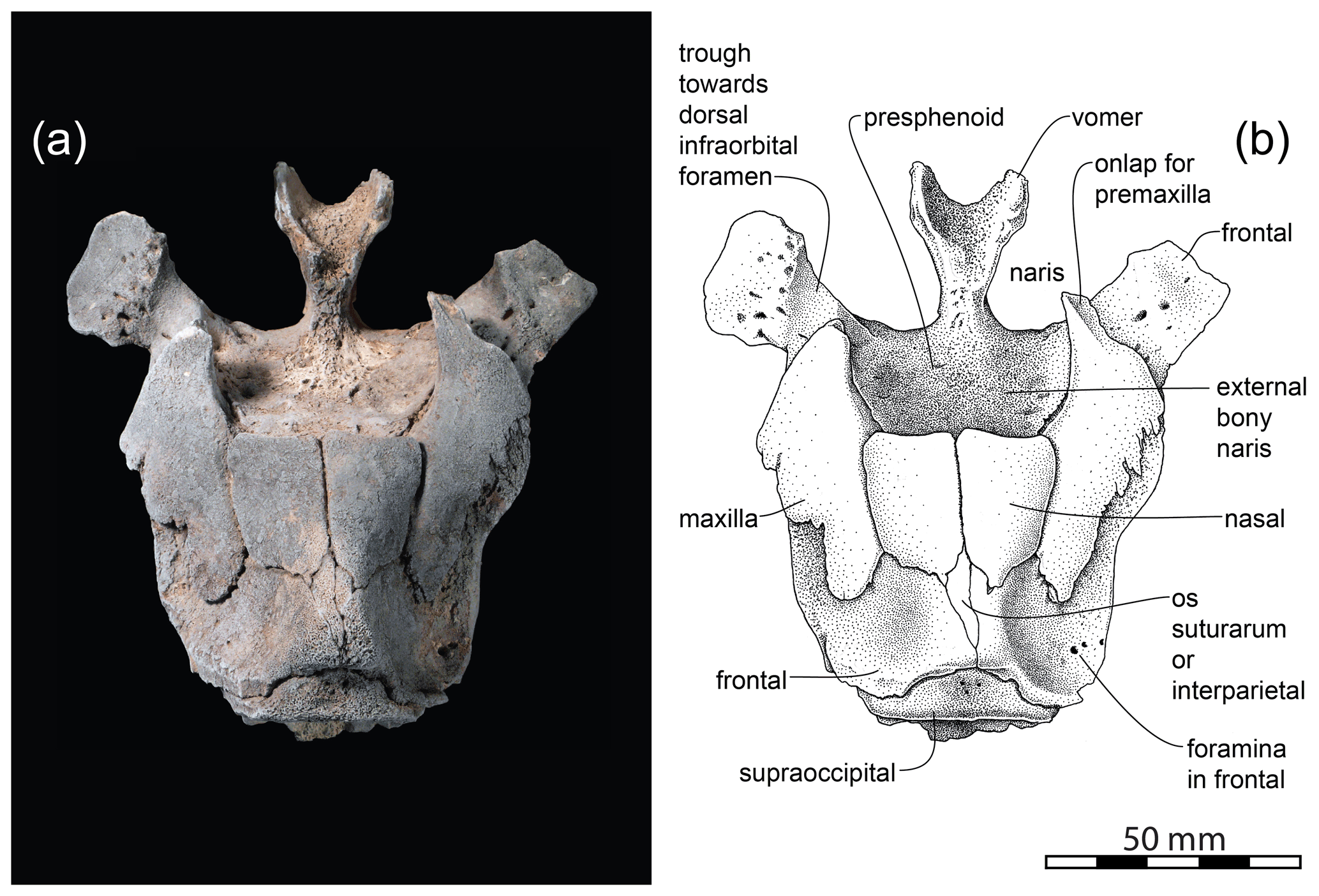
Figure 2Isoninia borealis (gen. nov., sp. nov.). (a) Partial skull CMM-V-4061 in dorsal view lightly coated with sublimed ammonium chloride. (b) Interpretative drawing of (a).
Vomer: the anterior-most preserved part of the skull is the prenarial process of the vomer (Figs. 2 and 3), which widens conspicuously to the mesorostral canal. Posterior to it is the nasal septum.
Maxilla: only parts of the maxillae adjacent to the nares and nasals are preserved. Here, at least their lateral margins are incomplete. However, with the preserved portion it can be clearly observed that the maxillae are retracted anteriorly such that they do not overlap the frontals immediately adjacent to the vertex (Figs. 2 and 3). Consequently, there is wide exposure of the frontal lateral to the vertex. Deep interdigitating sutures bind the maxillae and frontals (Fig. 3c). A shallow trough on the anterior extremity of the preserved segment of the right maxilla is interpreted as the area that held the posterior extremity of the now-missing premaxilla (Fig. 2). This suture indicates that the premaxillae were retracted rostrally such that the posterior-most reach of the premaxillae was approximately level with the mid-point in the length of the external bony nares. The maxillae form the posterolateral margin of the external nares. The thickened posteromedial margin of the maxilla broadly contacts and underlies the lateral margin of the nasal (Fig. 3a and c).
Nasals: both nasals are well preserved in Isoninia borealis. They are robust elements, each sub-rectangular in outline in both dorsal and lateral views. In dorsal view, their conjoined anterior margin forms the essentially straight posterior margin of the external bony nares. From this position, their combined widest point, they slightly narrow gradually along their lateral borders before turning abruptly medially towards the apex of the vertex. The dorsal surface of the nasals slopes conspicuously towards the bony nares (Fig. 3e). The pronounced undercut below the anterior margin of the nasals (Fig. 3c) may have held the now-missing part of the presphenoid that formed the posterodorsal margin of the nasal septum. The lateral margin of each nasal is deeply imbedded ventromedially into the frontal (Fig. 3c). The frontals are wedged between the posteromedial ends of the nasals.
Frontal: the frontals are the largest bones preserved in Isoninia borealis, occupying most of the dorsal area from the temporo-orbital plate to the back of the vertex (Figs. 2 and 3). The posterodorsal margin of the apex of the vertex may have been slightly eroded postmortem but is otherwise complete. Immediately medial to the preserved lateral margin of the frontal is a conspicuous trough until the margin of the preserved portion of the specimen, interpreted here as homologous to the trough for the posterior-most dorsal infraorbital foramen (the actual foramen is not preserved due to the loss of the lateral portion of the maxilla).

Figure 3Isoninia borealis (gen. nov., sp. nov.). (a) CMM-V-4061 in dorsal view; anterior to top of figure. (b) Single CT-scan image through CMM-V-4061 in posterior view as indicated by the transverse white line in (a) to which the corresponding arrow is pointing. (c) Single CT-scan image in posterior view through the anterior part of the nasals as indicated by the white line in (a). (d) Single CT-scan image in posterior view through the vertex showing the os suturarum or a fold in the frontals as indicated by the white line in (a). (e) CMM-V-4061 in left lateral view. (f) CMM-V-4061 in ventral view; anterior to top of figure. Specimen lightly coated with sublimed ammonium chloride. CT-scan images (b–d) are adjusted to the scale bar to the left below (a). The scale bar for (e) and (f) is to the left below (f).
The trough is (dorsal infraorbital canal; Fig. 3f) larger than in any other comparably sized odontocete, so the opening for the posterior-most dorsal infraorbital foramen was also thought to be correspondingly large, most similar to that seen in Isthminia (see Pyenson et al., 2015; fig. 3B). Immediately below this trough, ventrolaterally, a fossa for the postorbital lobe of the pterygoid sinus occupies the area on the ventrolateral face of the frontal below and behind the postorbital process of the frontal, (i.e., the postorbital recess in Fig. 3f).
Behind the thickened maxilla, the frontal gains wide dorsolateral exposure as it ascends the lateral face of the elevated vertex. The frontals form the narrow and pointed vertex posteromedially. Although the suture between contralateral frontals is located along the sagittal plane, below the vertex they are not symmetrical. A CT-scan image through the vertex transversely (Fig. 3d) confirms that the right frontal is thicker than the left and its lateral face is not as concave as is that of its left antimere. A supernumerary ossification is present on the vertex, being similar to structures described in the delphinid Sotalia guianensis (Simões-Lopes, 2006) as os suturarum. It is visible both in photos of the partial skull and in CT scans (Figs. 2, 3a and d). The CT images appear to indicate that it is a separate ossification distinct from the frontals, but lack of clarity engenders caution; it may simply be a fold in the left frontal. This separate ossification could represent an interparietal also present in Kwanzacetus and Inia (Lambert et al., 2018), as well as Samaydelphis (Lambert et al., 2020).
Below the vertex beyond the reach of the maxillae is a cluster of foramina that pass posteriorly and sub-horizontally through the frontal into the body of the supraoccipital (Fig. 2). The course of at least one larger foramen on either side of the vertex can be followed through the supraoccipital where it turns medially towards the midline of the latter.
The ventral sides of the frontals preserve some of the endocranial cavity (Fig. 3f). The ethmoid foramen passes from the anterior face of the cranial cavity anterolaterally towards the orbit. The dorsal rim of the optic canal is preserved posteroventral to the ethmoid foramen (Fig. 3e and f).
Presphenoid and cribriform plate: most of the presphenoid (Ichishima, 2016) is preserved in Isoninia borealis (Figs. 2 and 3). On the posterior cerebral face of the cribriform plate dorsomedial to the ethmoid foramina are two small ovoid depressions, the long axis of which has an anteroventral–posterodorsal orientation. They may have held small olfactory bulbs (or their vestigial remnants) in life; in any case, these cavities are less well developed as compared to the tiny olfactory bulb cavities preserved in the platanistoid Araeodelphis natator (Godfrey et al., 2017). Extending from each of these depressions is a trough that merges into a single one dorsomedially. Minute foramina pass from these depressions and conjoining troughs through the cribriform plate to the nasal cavity (Fig. 3b and f). Nevertheless, it is not known whether these tiny foramina through the cribriform plate actually conducted olfactory axons to olfactory receptor tissue within the nasal cavity (see Ichishima, 2016, p. 6). Unlike MGUH VP 3338 (an unnamed platanistoid odontocete from the latest Oligocene of Denmark; Hoch, 2000) and Squalodon sp. (Godfrey, 2013), there is no development of crescentic foramina or ethmoturbinals on the anterior side of the ectethmoid in Isoninia. Therefore, Isoninia borealis seems to be more derived in the complete loss of these structures over the two aforementioned osmatic odontocetes. Although the derived condition in Isoninia does not pinpoint its geological age, it does suggest a younger Miocene or Pliocene age – a time when most, if not all odontocetes were anosmatic.
Supraoccipital: a robust supraoccipital is wedged deeply between and behind the frontals (Fig. 3). In dorsal view, the supraoccipital gains wide exposure behind the vertex (Fig. 3a). Posterodorsally, the top of the supraoccipital is a step down from the top of the vertex formed by the frontals (Fig. 3e). In a posterior view of Isoninia, the broken bone suggests the presence of an external occipital crest just below the top of the supraoccipital. A transverse CT scan image through the vertex (Fig. 3d) shows that the supraoccipital was wedged dorsally between the thickened frontals. In ventral view, the supraoccipital abuts the frontals along their posteromedial margins. Posteriorly, within the brain cavity, the medial part of the supraoccipital becomes increasingly ridge-like (i.e., the internal occipital protuberance; Fig. 3d and f) and in life is presumed to have partially separated the cerebral hemispheres posteriorly.
In an attempt to test the phylogenetic position of Isoninia within modern and extinct odontocetes, we coded the specimen and conducted three experimental phylogenetic analyses using the morphological character matrix employed by Pyenson et al. (2015) and Lambert et al. (2020). In every analysis, the results differed widely, and Bremer support was very low. This kind of volatility and low support are to be expected when the analyses include fragmentary/poorly known taxa, (see also Post et al., 2017, and Lambert et al., 2018, 2020, for comments to this effect). Only when a more complete specimen of Isoninia with ear bones is found will the results of a phylogenetic analysis be trustworthy. Consequently, we have opted not to present a potentially misleading phylogenetic analysis here (however, the results of our analyses are presented in the Supplement).
Comparisons
Lambert et al. (2017) found that Inioidea is only diagnosed by a single unequivocal synapomorphy: width across nasals and nares subequal. Isoninia shares this feature with other inioids and forms the basis for our placement of Isoninia within this clade. Of the 14 equivocal synapomorphies for Inioidea, unfortunately because the holotype skull of Isoninia is so incomplete, it only preserves one – maxilla between premaxilla and nares edge.
We now compare Isoninia with some other inioids (listed alphabetically). Isoninia shares anteriorly retracted premaxillae with Auroracetus. Auroracetus differs from Isoninia in that its nasals appear to be more slender bilaterally than they are in Isoninia. Auroracetus is from the Nutrien Aurora Phosphate Mine in Aurora, North Carolina, USA. It derives from the Sunken Meadow Member of the Yorktown Formation and is therefore early Pliocene in age (Gibson and Geisler, 2009).
Isoninia shares the following features with Brachydelphis: nasals that narrow posteriorly and frontals wedged anteromedially between the nasals. Brachydelphis differs from Isoninia in that its frontals are only narrowly exposed on the vertex and the supraoccipital is not wedged forward between the frontals. Furthermore, in Brachydelphis, the premaxillae contact the nasals (although there is some intrageneric variation of that feature in Brachydelphis jahuayensis; see Lambert and Muizon, 2013), and the maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia. Two species of Brachydelphis are known (mazeasi and jahuayensis), both from Miocene localities in Chile and Peru (Gutstein et al., 2009; Lambert and Muizon, 2013).
Isoninia shares the following features with Brujadelphis: a moderately elevated vertex with wide nasals anteriorly that taper posterodorsally. Brujadelphis differs from Isoninia in that its frontals are very narrowly exposed on the vertex, it does not have a supraoccipital that is wedged between the frontals, nor does it have an interparietal, and the premaxillae and maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia. Brujadelphis is from the late Miocene Pisco Formation in Peru (Lambert et al., 2017).
Isoninia shares the following features with Inia: an elevated vertex in which the frontals are wedged between the posterior margins of the nasals, possibly an interparietal (Lambert et al., 2018), and a dorsomedial margin of the supraoccipital that is positioned between the frontals behind the vertex. The vertex of Inia differs from that of Isoninia in that the premaxillae and maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia,, and the vertex is not so abruptly elevated and does not show a frontal boss as seen in Inia. Furthermore, the nasals in Inia have a reduced dorsal exposure due to their position on the elevated anterior wall of the frontals on the vertex.
Isoninia shares the following features with Ischyrorhynchus: its premaxillae are retracted anteriorly so that they do not contact the nasals, and it possesses an elevated vertex, the top of which is formed by the frontals, and nasals that slope anteroventrally towards the external bony nares. Isoninia differs from Ischyrorhynchus in that its nasals are robust by comparison, and its frontals on the vertex are bilaterally compressed as opposed to being much wider and wider than long in Ischyrorhynchus. Ischyrorhynchus is known from the Miocene of Argentina, Brazil, and Venezuela (Pilleri and Gihr, 1979).
Isoninia shares the following features with Isthminia: a moderately elevated vertex in which the nasals taper posterodorsally, premaxillae and maxillae that are retraced anteriorly, and a large posterior dorsal infraorbital foramen. Isthminia differs from Isoninia in that the dorsal surface of the nasal slopes less steeply anteroventrally, and its right premaxilla appears to be longer posteriorly. Furthermore, the medial portion of the nasals is more elevated in Isthminia compared to the lateral part, and it does not seemingly have a supraoccipital that is wedged between the frontals. Isthminia is from the late Miocene Chagres Formation of Panama (Pyenson et al., 2015).
Isoninia shares the following features with Kwanzacetus: nasals that narrow posteriorly and an elevated vertex, possibly also preserving an interparietal. Kwanzacetus has a medially and undercut abruptly elevated vertex as in other iniids, which is not observed in Isoninia, that has a more gradually sloped and not so elevated vertex. The frontal boss present in Kwanzacetus, as in Inia and Ischyrorhynchus, is not present in Isoninia. The right premaxilla is also most likely longer posteriorly in Kwanzacetus. Furthermore, the lateral margin of the nasal is much more rounded, and there is a shallow internasal fossa in Kwanzacetus. Kwanzacetus is known from the late Miocene Kwanza Basin of Angola (Lambert et al., 2018).
Isoninia shares the following features with Meherrinia: a smooth and moderately elevated vertex, in which the frontals are wedged between the posterior margins of the nasals, and a supraoccipital that is also wedged between the posterior margins of the frontals. The vertex of Meherrinia differs from that of Isoninia in that it is proportionately narrower, the nasals are not pinched posteriorly, and the premaxillae and maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia. Meherrinia is probably from the late Miocene Eastover Formation in North Carolina, USA (Geisler et al., 2012).
Isoninia shares the following features with Pliopontos: frontals that are wedged anteromedially between the nasals and a supraoccipital that is wedged between the posteromedial margins of the frontals. Pliopontos differs from Isoninia in that it does not have as elevated a vertex, wide nasals throughout their length, and premaxillae and maxillae that are not retracted anteriorly to the same degree. In Pliopontos, the nasals are proportionately elongated, extending posteriorly almost making contact with the supraoccipital (de Muizon, 1984; fig. 1). Consequently, the frontals gain only very limited exposure on the vertex, to which the nasals rise to the same level as the frontals. Pliopontos derives from the Pliocene Pisco Formation of Peru (de Muizon, 1983).
Pontistes differs from Isoninia in that it has premaxillae that contact the nasals, a low cranial vertex, maxillae that extend posteriorly to the level of the supraoccipital, and a supraoccipital that does not wedge deeply between the frontals. Pontistes is from the Miocene Entrerriana Formation of Argentina (Cozzuol, 2010).
Isoninia shares the following features with Pontoporia: nasals that taper posteriorly, frontals that are wedged between the nasals posteriorly, and a supraoccipital that is wedged between the poster margins of the frontals. (However, in one skull of Pontoporia that Stephen J. Godfrey has seen, the posterior margins of the nasals are wedged between the frontals.) The vertex of Pontoporia differs from that of Isoninia in that it is much lower, and the premaxillae and maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia.
Isoninia shares the following features with Samaydelphis: a moderately elevated vertex, in which the nasals become narrower posteriorly, frontals that are wedged between the nasals, and possibly a dorsally exposed interparietal. The vertex of Samaydelphis differs from that of Isoninia in that the nasals rise to the same level as the frontals, the frontals are more deeply wedged between the nasals than in Isoninia, and the premaxillae and maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia. Samaydelphis originates from within the late Miocene marine deposits of the Pisco Formation of the East Pisco Basin, Peru (Lambert et al., 2020).
Isoninia shares the following features with Scaldiporia: a moderately elevated vertex and nasals that narrow posteriorly. Scaldiporia differs from Isoninia in that it has an internasal fossa, its frontals are very narrowly exposed on the vertex, it does not seemingly have a supraoccipital that is wedged between the frontals, nor does it have an interparietal, and the premaxillae and maxillae are not retracted anteriorly to the same extent as they appear to be in Isoninia. Scaldiporia is from marine deposits of the Breda Formation – Late Miocene to earliest Pliocene of the Westerschelde estuary (the Netherlands) (Post et al., 2017).
Isoninia shares the following features with Stenasodelphis: a moderately elevated vertex, in which the nasals become narrower posteriorly, frontals that are wedged between the nasals, and possibly a shared dorsally exposed interparietal. Stenasodelphis differs from Isoninia in that its premaxillae and maxillae are not retracted anteriorly to the same degree. Stenasodelphis probably derives from the late Miocene Little Cove Point Member of the St. Marys Formation in Maryland, USA (Godfrey and Barnes, 2008).
In most of the aforementioned inioids, the posterior ends of the premaxillae are retracted anteriorly so that they do not contact the nasals (and consequently, the premaxillae are widely separated from the posterior margin of the external bony nares). This feature characterizes all inioids (and many extant delphinoids) except Pontistes rectifrons (Burmeister, 1885; de Muizon, 1983, 1984) and Brachydelphis (Gutstein et al., 2009, although it is variable). Isoninia differs from other inioids in that its nasals are robust. The development of an incipient frontal boss in Isoninia and that its supraoccipital is deeply wedged forward between the frontals suggest the placement of this new species within the family Iniidae.
North American inioids include Auroracetus bakerae Gibson and Geisler, 2009; Goniodelphis hudsoni Allen, 1941; Meherrinia isoni Geisler, Godfrey, and Lambert 2012; Stenasodelphis russellae Godfrey and Barnes, 2008; and Isoninia borealis. All of these taxa are poorly known. Assigning Isoninia to the Inioidea increases the morphological diversity exhibited by this clade during the late Miocene and Pliocene outside South America.
Although Isoninia borealis presents a combination of features that warrant the naming of a new genus and species within the Odontoceti, unfortunately, because the holotype is incomplete, phylogenetic analyses are highly volatile and very weekly supported (see Supplement). Nevertheless, describing another Miocene inioid from North America, as suggested by the shared characters established through comparisons to other inioids, further testifies to the morphological diversity found in this clade.
The present new record from the Atlantic Coastal Plain of the United States, together with inioid genera found in Angola (Lambert et al., 2018), the Caribbean (Cozzuol and Aguilera, 2008; Pyenson et al., 2015; Aguirre-Fernandez et al., 2017), Japan (Murakami, 2016), the North Sea (Lambert and Post, 2005; Pyenson and Hoch, 2007; Post et al., 2017), and Peru (Lambert et al., 2017, 2020), reinforces the hypothesis that extant inioids have a relictual distribution (Cassens et al., 2000; Gutstein et al., 2014; Pyenson et al., 2015).
All data and results are presented in the main text, tables, figures, and the Supplement.
The supplement related to this article is available online at: https://doi.org/10.5194/fr-24-275-2021-supplement.
SJG prepared and edited the manuscript and figures. CSG prepared and edited the manuscript. DJM III undertook the experimental phylogenetic analyses and edited the manuscript.
The authors declare that they have no conflict of interest.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
We extend our gratitude to Ronald Ison for scuba-collecting and donating the specimen described herein to the Calvert Marine Museum. The murky, fallen-tree entangled, riverine conditions in which this specimen was found were anything but ideal. We would also like to thank Bruno Frohlich (NMNH) for CT scanning the holotype specimen. John R. Nance (CMM), Nicholas Pyenson (NMNH), David J. Bohaska (NMNH), John Ososky (NMNH), and Charles W. Potter (retired from the NMNH) are gratefully acknowledged for encouraging the study of fossil and modern odontocetes in their care. The late John Pojeta (NMNH) welcomed us to his lab where CMM-V-4061 was whitened with sublimed ammonium chloride.
Alberto Collareta and Olivier Lambert provided very constructive reviews. For detailed reviews of an earlier version of this article, we are grateful to Gabriel Aguirre Fernández and Mary T. Silcox.
This publication was made possible with funding for Stephen J. Godfrey and Donald J. Morgan III from the Citizens of Calvert County, the County Board of Calvert County Commissioners, MD, and the Clarissa and Lincoln Dryden Endowment for Paleontology at the Calvert Marine Museum. Funding for Carolina S. Gutstein came from her Postdoctoral Fondecyt no. 3160710, ACT 172099 PIA-Conicyt (PI: Alexander O. Vargas), and REDES190190 (PI: Alexander O. Vargas).
This paper was edited by Florian Witzmann and reviewed by Olivier Lambert and Alberto Collareta.
Aguirre-Fernández, G., Carrillo-Briceño, J. D., Sánchez, R., Amson, E., Sánchez-Villagra, M. R.: Fossil cetaceans (Mammalia, Cetacea) from the Neogene of Colombia and Venezuela, J. Mamm. Evol., 24, 71–90, https://doi.org/10.1007/s10914-016-9353-x, 2017.
Allen, G.: A fossil river dolphin from Florida, Bulletin of the Museum of Comparative Zoology, 89, 3–11, 1941.
Ameghino, F.: Mamiferos y aves fósiles Argentinas: Especies nuevas, adiciones, y correciones, Revista Argentina de Historia Natural, 1, 240–259, 1891.
Arnason, U. and Gullberg, A.: Cytochrome bnucleotide sequences and the identification of five primary lineages of extant cetaceans, Mol. Biol. Evol., 13, 407–417, 1996.
Banguera-Hinestroza, E., Cardenas, H., Ruiz-García, M., García, Y. F., Marmontel, M., Gaitán, E., Vásquex, R., and García-Vallejo, F.: Molecular identification of evolutionarily significant units in the Amazon River Dolphin Inia sp. (Cetacean Iniidae), J. Hered., 93, 321–322, 2002.
Brisson, M. J.: Le regnum animale in classes IX distributum, sive synopsis methodica sistens generalem animalium distributionem in classes IX, & duarum primarum classium, quadrupedum scilicet & cetaceorum, particularem divisionem in ordines, sectiones, genera & species, T. Haak, Paris, 296 pp., 1762.
Burmeister, G.: On Saurocetes argentinus, a new type of Zeuglodontidae, Ann. Mag. Nat. Hist., 7, 51–55, 1871.
Burmeister, G.: Examen critico de los mamíferos y reptiles fósiles denominados por D. Augusto Bravard y mencionados en su obra precedente. Anales del Museo Nacional de Buenos Aires, Tomo 3 Serie I (entrega XIV), 93–174, 1885.
Cassens, I., Vicario, S., Wadde, V. G., Balchowsky, H., Balle, D. V., Wang, D., Chen, F., Mohan, R. S. L., Simoes-Lopes, P. C., Bastida, R., Meyer, A., Stanhope, M. J., and Millinkovitch, M. C.: Independent adaptation to riverine habitats allowed survival of ancient cetacean lineages, P. Natl. Acad. Sci. USA, 97, 11343–11347, 2000.
Cione, A. L., Cozzuol, M. A., Dozo, M. T., and Acosta-Hospitaleche, C.: Marine vertebrate assemblages in the southwest Atlantic during the Miocene, Biol. J. Linn. Soc., 103, 423–440, 2011.
Cooper, C. L.: Ammonium chloride sublimate apparatus, J. Paleontol., 9, 357–359, 1935.
Cozzuol, M. A.: The Odontoceti of the “Mesopotamiense” of the Paraná River ravines, Systematic review, Investigations on Cetacea, 7, 39–53, 1985.
Cozzuol, M. A.: Una nueva especie de Saurodelphis Burmeister, 1891 (Cetacea: Iniidae) del “Mesopotamiense” (Mioceno tardío-Plioceno temprano) de la provincia de Entre Ríos, Argentina, Ameghiniana, 25, 39–45, 1989.
Cozzuol, M. A.: The record of the aquatic mammals in southern South America, Münchner Geowissenschaftliche Abhandlungen, Reihe A, Geologie und Paläontologie, 30, 321–342, 1996.
Cozzuol, M. A.: Fossil record and the evolutionary history of Iniodea, in: Biology, Evolution and Conservation of River Dolphins within South America and Asia, edited by: Ruiz-García, M. and Shostell, J., 193–217, Nova Science Publishers, New York, 2010.
Cozzuol, M. A. and Aguilera, O. A.: Cetacean remains from the Neogene of northwestern Venezuela, Paläontol. Z., 82, 196–203, 2008.
Cremer, M. J. and Simões-Lopes, P. C.: The occurrence of Pontoporia blainvillei (Gervais & d'Orbigny) (Cetacea, Pontoporiidae) in an estuarine area in southern Brazil, Rev. Bras. Zool., 22, 717–723, 2005.
de Blainville, H.: Nouveau dictionnaire d'histoire naturelle, edited by: Desmarest, A. G., Vol. 9, 151 pp., 1817.
de Muizon, C.: Pliopontos littoralis un nouveau Platanistidae (Cetacea) du Pliocène de la côte péruvienne, Comptes Rendus de l'Académie des Sciences Paris, Série II, 296, 1101–1104, 1983.
de Muizon, C.: Les vertébrés fossiles de la Formation Pisco (Pérou). Deuxième partie: les odontocètes (Cetacea, Mammalia) du Pliocène inférieur de Sud-Sacaco, Institut Francais d'Études Andines, Mémoire, 50, 1–188, 1984.
de Muizon, C.: Les relations phylogénétiques des Delphinida (Cetacea; Mammalia), Annales de Paléontologie (Vert.-Invert.), Editions Recherche sur les civilizations, 74, 159–227, 1988.
Feldman, R. M.: Whitening fossils for photographic purposes, in: Paleotechniques, edited by: Feldman, R. M., The Paleontological Society Special Publication, 4, 342–346, 1989.
Flower, W. H.: Description of the skeleton of Inia geoffrensis and of the skull of Pontoporia blainvillei, with remarks on the systematic position of these animals in the order Cetacea, Transactions of the Zoological Society of London, 6, 87–116, 1867.
Gatesy, J., Geisler, J. H., Chang, J., Buell, C., Berta, A., Meredith, R. W., Springer, M. S., and McGowen M. R.: A phylogenetic blueprint for a modern whale, Mol. Phylogenet. Evol., 66, 479–506, 2013.
Geisler, J. H. and Sanders, A. E.: Morphological evidence for the phylogeny of Cetacea, J. Mamm. Evol., 10, 23–129, 2003.
Geisler, J. H., McGowen, M. R., Yang, G., and Gatesy, J.: A supermatrix analysis of genomic, morphological, and paleontological data for crown Cetacea, BMC Evol. Biol., 11, 1–22, 2011.
Geisler, J. H., Godfrey, S. J., and Lambert, O.: A new genus and species of Late Miocene inioid (Cetacea: Odontoceti) from the Meherrin River, North Carolina, U.S.A., J. Vertebr. Paleontol., 32, 198–211, 2012.
Gervais, F. L. P. and D'orbigny, A.: M. Paul Gervais, au nom de M. Alcide D'Orbigny et au sien, met sous les yeux de la Société trois planches représentant des Dauphins observés par ce dernier pendant son voyage dans l'Amérique méridionale, Bulletin de la Société Philomathique de Paris, 27 April, 38–40, 1844.
Gibson, M. L. and Geisler, J. H.: A new Pliocene dolphin (Cetacea: Pontoporiidae), from the Lee Creek Mine, North Carolina, J. Vertebr. Paleontol., 29, 966–971, 2009.
Gill, T.: Synopsis of the primary subdivisions of the cetaceans, Communications of the Essex Institute, 6, 121–126, 1871.
Godfrey, S. J.: On the olfactory apparatus in the Miocene odontocete Squalodon sp. (Squalodontidae), C.R. Palevol., 12, 519–530, 2013.
Godfrey, S. J. and Barnes, L. G.: A new genus and species of Late Miocene pontoporiid dolphin (Cetacea: Odontoceti) from the St. Marys Formation in Maryland, J. Vertebr. Paleontol., 28, 520–528, 2008.
Godfrey, S. J., Barnes, L. G., and Lambert, O.: The identity of the Early Miocene dolphin Araeodelphis natator Kellogg, 1957 (Cetacea; Platanistidae), from the Calvert Formation, J. Vertebr. Paleontol., 37, e1278607, https://doi.org/10.1080/02724634.2017.1278607, 2017.
Gray, J. E.: On the cetaceous animals, The zoology of the voyage of H. M. S. Erebus and Terror, under the command of Capt. Sir J.C. Ross, R.N., F.R.S., during the years 1839 to 1843 1 and 2, 1–53, E.W. Janson, London, 1846.
Gutstein, C. S., Cozzuol, M. A., Vargas, A. O., Suárez, M. E., Schultz, C. L., and Rubilar-Rogers, D.: Patterns of skull variation of Brachydelphis (Cetacea, Odontoceti) from the Neogene of the southeastern Pacific, J. Mammal., 90, 504–519, 2009.
Gutstein, C. S., Cozzuol, M. A., and Pyenson, N. D.: The antiquity of riverine adaptations in Iniidae (Cetacea, Odontoceti) documented by a humerus from the Late Miocene of the Ituzaingó Formation, Argentina, Anat. Rec., 297, 1096–1102, 2014.
Hamilton, H., Caballero, S., Collins, A. G., and Brownell Jr., R. L.: Evolution of river dolphins, Royal Society, 268, 549–556, 2001.
Hoch, E.: Olfaction in whales: evidence from a young odontocete of the late Oligocene north sea, Hist. Biol., 14, 67–89, 2000.
Hrbek, T., Da Silva, V. M. F., Dutra, N., Gravena, W., Martin, A. R., and Farias, I. P.: A new species of river dolphin from Brazil or: How little do we know our biodiversity, PLoS ONE, 9, e83623, https://doi.org/10.1371/journal.pone.0083623, 2014.
Ichishima, H.: The ethmoid and presphenoid of cetaceans, J. Morphol., 277, 1661–1674, https://doi.org/10.1002/jmor.20615, 2016.
Lambert, O. and Post, K.: First European pontoporiid dolphins (Mammalia: Cetacea, Odontoceti), from the Miocene of Belgium and The Netherlands, Deinsea, 11, 7–20, 2005.
Lambert, O. and de Muizon, C.: A new long-snouted species of the Miocene pontoporiid dolphin Brachydelphis and a review of the Mio-Pliocene marine mammal levels in the Sacaco Basin, Peru, Jo. Vertebr. Paleontol., 33, 709–721, 2013.
Lambert, O., Bianucci, G., Urbina, M., and Geisler, J. H.: A new inioid (Cetacea, Odontoceti, Delphinida) from the Miocene of Peru and the origin of modern dolphin and porpoise families, Zool. J. Linn. Soc., 179, 919–946, 2017.
Lambert, O., Auclair, C., Cauxeiro, C., Lopez, M., and Adnet, S.: A close relative of the Amazon river dolphin in marine deposits: a new Iniidae from the late Miocene of Angola, 6, e5556, https://doi.org/10.7717/peerj.5556, 2018.
Lambert, O., Collareta, A., Benites-Palomino, A., Di Celma, C., De Muizon, C., Urbina, M., and Bianucci, G.: A new small, mesorostrine inioid (Cetacea, Odontoceti, Delphinida) from four upper Miocene localities in the Pisco Basin, Peru, Papers in Paleontology, 7, 1043–1064, https://doi.org/10.1002/spp2.1332, 2020.
McGowen, M. R., Spaulding, M., and Gatesy, J.: Divergence date estimation and a comprehensive molecular tree of extant cetaceans, Mol. Phylogenet. Evol., 53, 891–906, 2009.
McGowen, M. R., Tsagkogeorga, G., Álvarez-Carretero, S., dos Reis, M., Struebig, M., Deaville, R., Jepson, P. D., Jarman, S., Polanowski, A., and Morin, P. A.: Phylogenomic resolution of the cetacean tree of life using target sequence capture, Syst. Biol., 69, 479–501, 2020.
Mead, J. G. and Fordyce, R. E.: The therian skull: a lexicon with emphasis on the odontocetes, Smithsonian Contributions to Zoology, 627, 1–248, 2009.
Morgan, G. S.: Miocene and Pliocene marine mammal faunas from the Bone Valley Formation of central Florida, in: Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore, Jr., edited by: Berta, A. and Deméré, T. A., Proceedings of the San Diego Society of Natural History, 29, 239–268, 1994.
Murakami, M.: A New Extinct Inioid (Cetacea, Odontoceti) from the Upper Miocene Senhata Formation, Chiba, Central Japan: The First Record of Inioidea from the North Pacific Ocean, Paleontol. Res., 20, 207–225, https://doi.org/10.2517/2015PR031, 2016.
Nikaido, M., Matsuno, F., Hamilton, H., Brownell Jr., R. L., Ying, C., Wang, D., Zhu, Z. Y., Shedlock, A. M., Fordyce, R. E., Hasegawa, M., and Okada, N.: Retroposon analysis of major cetacean lineages: the monophyly of the toothed whales and the paraphyly of river dolphins, P. Natl. Acad. Sci. USA, 98, 7384–7389, 2001.
Pilleri, G. and Gihr, M.: Skull, sonar field and swimming behavior of Ischyrorhynchus vanbenedeni (Ameghino, 1891) and taxonomical position of the genera Ischyrorhynchus, Saurodelphis, Anisodelphis and Pontoplanodes (Cetacea), Investigations on Cetacea, 5, 17–69, 1979.
Post, K., Louwye, S., and Lambert, O.: Scaldiporia vandokkumi, a new pontoporiid (Mammalia, Cetacea, Odontoceti) from the Late Miocene to earliest Pliocene of the Westerschelde estuary (The Netherlands), PeerJ, 5, e3991, https://doi.org/10.7717/peerj.3991, 2017.
Pyenson, N. D. and Hoch, E.: Tortonian pontoporiid odontocetes from the Eastern North Sea, J. Vertebr. Paleontol., 27, 757–762, 2007.
Pyenson, N. D., Velez-Juarbe, J., Gutstein, C. S., Little, H., Vigil, D., and O'dea, A.: Isthminia panamensis, a new fossil inioid (Mammalia, Cetacea) from the Chagres Formation of Panama and the evolution of “river dolphins” in the Americas, PeerJ, 3, e1227, https://doi.org/10.7717/peerj.1227, 2015.
Ruiz-Garcia, M. and Shostell, J. M.: Biology, evolution and conservation of river dolphins within South America and Asia, Nova Science Publishers, Hauppauge, 1–504, 2010.
Secchi, E. R.: Life history and ecology of Franciscana, Pontoporia blainvillei (Cetacea, Pontoporiidae), in: Biology, Evolution and Conservation of River Dolphins within South America and Asia, edited by: Ruiz-García, M. and Shostell, J., 301–322, Nova Science Publishers, New York, 2010.
Shelburne, E. C. H. and Thompson, A. C.: Specimen whitening: An assessment of methods of ammonium chloride smoke removal, Collection Forum, 30, 63–72, 2016.
Simões-Lopes, P. C.: Morfologia do sincrânio do boto-cinza, Sotalia guianensis (P.J. van Bénéden) (Cetacea, Delphinidae), Rev. Bras. Zool., 23, 652–660, 2006.
Ward, L. W. and Andrews, G. W.: Stratigraphy of the Calvert, Choptank, and St. Marys Formations (Miocene) in the Chesapeake Bay area, Maryland and Virginia, Virginia Museum of Natural History, Memoir Number 9, 60 pp., 2008.
Whitmore Jr., F. C. and Kaltenbach, J. A.: Neogene Cetacea of the Lee Creek Phosphate Mine, North Carolina, Virginia Museum of Natural History Special Publication, 14, 181–269, 2008.
Yang, G. and Zhou, K.: A study on the molecular phylogeny of river dolphins, Acta Theriologica Sinica, 19, 1–9, 1999.
Yang, G., Yan, J., Zhou, K., and Wei, F.: Sequence variation and gene duplication at MHC DQB Loci of Baiji (Lipotes vexillifer), a Chinese River Dolphin, J. Hered., 96, 310–317, https://doi.org/10.1093/jhered/esi055, 2005.