the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 12 Feb 2020
| 12 Feb 2020
A new species of baenid turtle from the Early Cretaceous Lakota Formation of South Dakota
Walter G. Joyce
Yann Rollot
Richard L. Cifelli
Baenidae is a clade of paracryptodiran turtles known from the late Early Cretaceous to Eocene of North America. The proposed sister-group relationship of Baenidae to Pleurosternidae, a group of turtles known from sediments dated as early as the Late Jurassic, suggests a ghost lineage that crosses the early Early Cretaceous. We here document a new species of paracryptodiran turtle, Lakotemys australodakotensis gen. and sp. nov., from the Early Cretaceous (Berriasian to Valanginian) Lakota Formation of South Dakota based on a poorly preserved skull and two partial shells. Lakotemys australodakotensis is most readily distinguished from all other named Late Jurassic to Early Cretaceous paracryptodires by having a broad, baenid-like skull with expanded triturating surfaces and a finely textured shell with a large suprapygal I that laterally contacts peripheral X and XI and an irregularly shaped vertebral V that does not lap onto neural VIII and that forms two anterolateral processes that partially separate the vertebral IV from contacting pleural IV. A phylogenetic analysis suggests that Lakotemys australodakotensis is a baenid, thereby partially closing the previously noted gap in the fossil record.
- Article
(11747 KB) - Full-text XML
-
Supplement
(461 KB) - BibTeX
- EndNote




Baenidae is a clade of fossil paracryptodiran turtles known from the Early Cretaceous to Eocene of North America (Joyce and Lyson, 2015). Late Cretaceous to Eocene basins throughout the western United States and Alberta (e.g., Hay, 1908; Gaffney, 1972; Brinkman, 2003, 2005; Joyce and Lyson, 2015) have yielded rich fossil turtle material indicating that baenids were diverse throughout this time period and were only little affected by the K–T mass extinction event (e.g., Lyson et al., 2011, 2019). Only little is known, by contrast, about the early evolution of the group, mostly due to a lack of fossils. At present, only three unambiguous baenid fossils are known from the Early Cretaceous, in particular the holotype specimens of Arundelemys dardeni Lipka et al., 2006, Trinitichelys hiatti Gaffney, 1972, and Protobaena wyomingensis (Gilmore, 1919) from the Aptian–Albian of Maryland, Texas, and Wyoming, respectively. A broad set of mostly unfigured fragmentary remains from the Early Cretaceous of Nevada (Bonde et al., 2008), Montana (Oreska et al., 2013), Oklahoma (e.g., Cifelli et al., 1997), and Utah (e.g., Cifelli et al., 1999) may represent fossil baenids as well, but are too incomplete to allow rigorous identification.
We present new fossil turtle material from the Early Cretaceous (Berriasian to Valanginian) Lakota Formation of South Dakota, which we identify as a new species of baenid, Lakotemys australodakotensis gen. and sp. nov. The purpose of this contribution is to describe this new material and to place it in a phylogenetic context.
The specimens described herein were collected from two localities in exposures of the Lakota Formation in the Cheyenne River drainage, between Devil and Hell canyons, Fall River County, South Dakota (Fig. 1). The localities, separated by about 1.5 km, lie on adjacent parts of the Flint Hill and Cascade Springs 7.5′ quadrangles, both of which have been geologically mapped (Post, 1967; Bell and Post, 1971). OMNH locality V1332 (sec. 34, T8S R4E; precise locality data are on file at OMNH and are available to qualified investigators upon request) lies within Dick Canyon, close to its mouth at the Cheyenne River. This site yielded a partial skull (OMNH 66106) and a nearly complete shell (OMNH 67133), described below. OMNH locality V1382 (sec. 3, T9S R4E), is on the north bank of the Cheyenne River below the mouth of Devil Canyon. This site yielded a second, nearly complete shell (OMNH 63615) described herein as well. Also present at V1382 but not collected because of logistic issues were several patches of articulated ganoid fish scales.
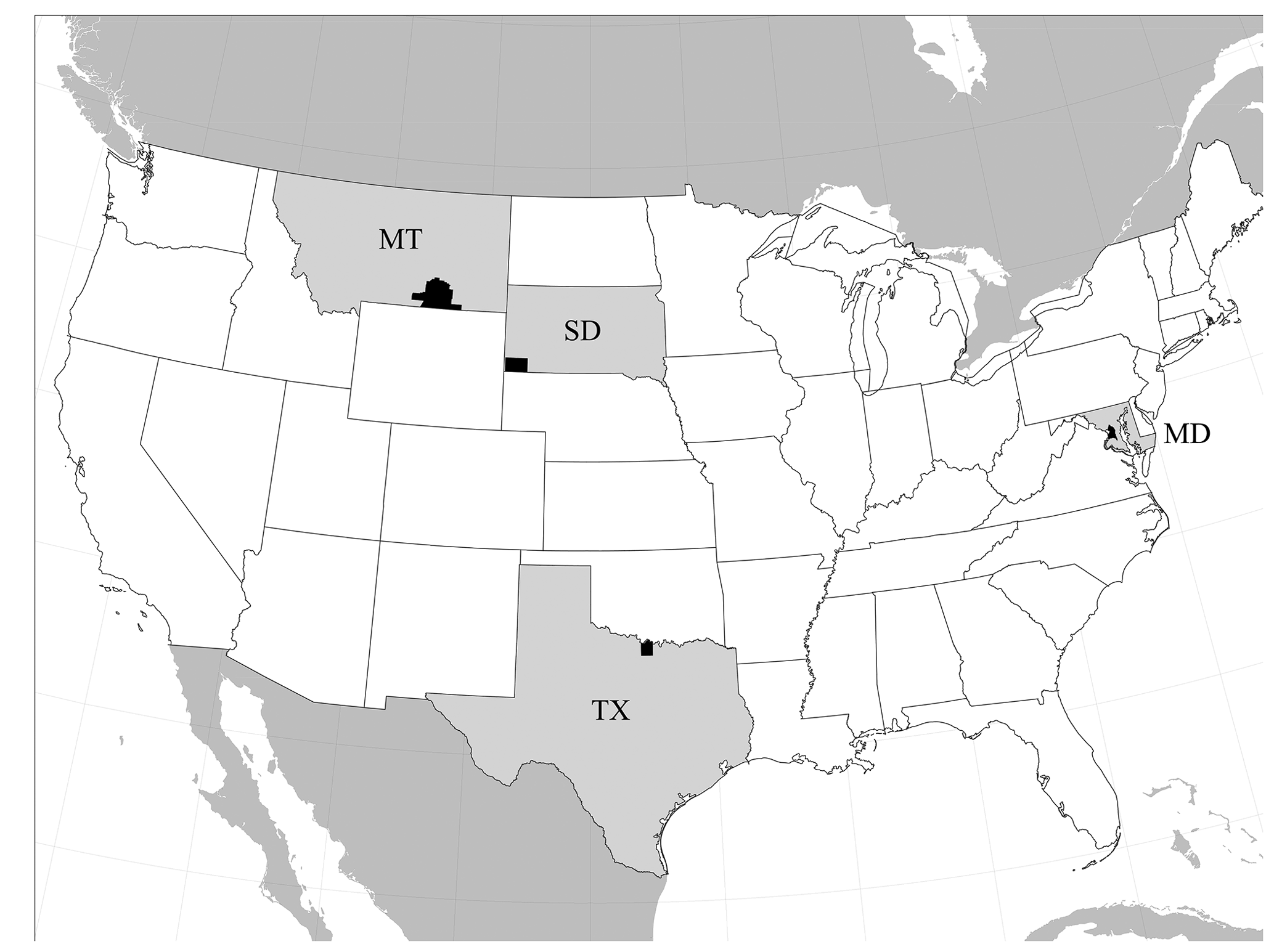
Figure 1Map highlighting the distribution of named baenids from the Early Cretaceous of North America: Arundelemys dardeni from St. George's County, Maryland (MD); Lakotemys australodakotensis gen. et. sp. nov. from Fall River County, South Dakota (SD); Protobaena wyomingensis from Big Horn County, Montana (MT); and Trinitichelys hiatii from Montague County, Texas (TX). States are highlighted in gray, counties in black.
The Lakota and overlying Fall River formations comprise the Inyan Kara Group, which crops out in an elliptical belt (Ward, 1894) around the periphery of the Black Hills Uplift in southwestern South Dakota and northeastern Wyoming (Waagé, 1959). The Inyan Kara Group is one of several rock units resulting from the accumulation of terrigenous to marginal marine sediments in the Western Interior foreland basin, east of the North American Cordilleran orogenic belt, during the Early Cretaceous (DeCelles, 2004). Most of these units, including the Lakota Formation, unconformably overlie the Upper Jurassic Morrison Formation. Locally, the Lakota rests unconformably on the Unkpapa Sandstone, an aeolian unit sometimes regarded as a localized facies of the upper Morrison (Szigeti and Fox, 1981).
The Lakota Formation is principally made up of massive, fluvial sandstones that interbed and intergrade laterally with paludal and lacustrine mudstones and limestone (Waagé, 1959; Bell and Post, 1971). A threefold subdivision into informal units, L1–3 (Way et al., 1998; Zaleha, 2006), or three named members (Waagé, 1959; Post and Bell, 1961) is generally employed for the Lakota Formation. Unfortunately, the subdivisions are not equivalent between the systems. The Chilson (lowest) and Minnewaste Limestone members are both placed in L1; the uppermost or Fuson Member includes both L2 and L3.
The Lakota Formation varies from 15 to 150 m thick and is thickest in the southern and southwestern part of the Black Hills uplift (Dahlstrom and Fox, 1995), which includes the study area (operationally defined herein as the area including the two localities and their immediate vicinity). Lithostratigraphic correlation between widespread areas can be difficult owing to considerable variation in bed thickness and composition (Bolyard and McGregor, 1966). Where the highly distinctive Minnewaste Limestone Member is absent, similar lithologies within the Chilson and Fuson members can lead to confusion in areas with limited, patchy exposure (Cifelli et al., 2014). However, the sandstones have been studied in some detail, and they provide an adequate basis for correlation in much of the Black Hills region. Four distinctive, large-scale fluvial units (distinctive sandstone bodies with associated mudstones), commonly designated (bottom to top) s1–4 (sandstones) and m1–4 (mudstones) on maps and measured sections, have been recognized by Post and Bell (1961) and subsequent workers (Post, 1967; Bell and Post, 1971; Gott et al., 1974; Dandavati, 1985; Dahlstrom and Fox, 1995). Fluvial units 1 and 2 occur in the Chilson Member and fluvial units 3 and 4 are placed in the Fuson Member. To the west of the study area, in Chilson Canyon, both fluvial complexes (or units) 1 and 2 are present in the Chilson Member, but in the immediate area of the localities, unit 1 is absent. At the mouth of Dick Canyon (Post, 1967), sandstone containing crocodilian and theropod tracks, and attributable to unit 2, lies at the canyon floor, followed up-section by about 20 m of gray mudstone with some minor interbedded sandstones. OMNH locality V1332 lies just above the base of a massive, unit 2 sandstone overlying the gray mudstone. This channel sandstone and its contact with the underlying gray mudstone can be traced up the canyon of the Cheyenne River. At a point adjacent to OMNH locality V1382, about 1500 m to the west, the contact occurs near low-water river level (Bell and Post, 1971), and the fossiliferous level is within the lowest part of the overlying massive sandstone. Hence, both turtle localities lie in the same stratigraphic horizon: a sandstone in the lower part of unit 2, Chilson Member, Lakota Formation.
Recognition of the Lakota Formation as a distinct lithologic unit (Darton, 1901) followed quickly on the heels of the discovery of cycads (characteristic of Early Cretaceous or older units) in the lower part of beds previously regarded as belonging to the Upper Cretaceous “Dakota” Group (Ward, 1894). Beyond the general determination of an Early Cretaceous age for the Lakota Formation, little progress was made for more than a century thereafter. Based on non-marine ostracods, Sohn (1979, and references cited therein) proposed that the Lakota was Valanginian–Barremian in age, far older than the Aptian age cited in many other studies (see Sames et al., 2010, and references cited therein). Zaleha (2006) pointed out that most of Sohn's samples had come from the lower part of the unit (Chilson and Minnewaste Limestone members, or L1 of Way et al., 1998) and correlated the upper part of the Lakota (Fuson Member, or L2–3 of Way et al., 1998) with Cloverly B and C intervals, considering them to be of early–middle Albian age. An Albian correlation for these parts of the Cloverly, at least, is upheld by radiometric determinations (D'Emic et al., 2019).
The most substantial biostratigraphic evidence for the age of the Lakota Formation comes from the study of existing and new ostracod collections, together with comparison to well-documented sequences from Europe (Sames et al., 2010; Sames, 2011). Based on these data, the unit ranges from Berriasian to Barremian, with respective, overlapping estimates of Berriasian–Valanginian and Valanginian–Barremian for the Chilson and Fuson members (Cifelli et al., 2014). Accepting these estimates for present purposes, we regard the turtle specimens described herein, from unit 2 of the Chilson Member, to be of Berriasian–Valanginian age, or about 133–140 Myr old, following the absolute ages of Walker et al. (2018).
High-resolution X-ray micro-computed tomography was applied to all available specimens at the University of Texas High-Resolution X-ray Computed Tomography Facility, USA, in June 2010. OMNH 63615 was scanned with 1000 projections over 360∘, a brass filter, an exposure time of 32 ms, a voltage of 450 kV, and a current of 3000 µA, which were converted into 609 coronal slices with a voxel size of 500 µm. OMNH 66106 was scanned with 1400 projections over 360∘, a voltage of 210 kV, and a current of 170 µA, which were converted into 1029 coronal slices with a voxel size of 59 µm. OMNH 67133 was scanned with 1000 projections over 360∘, a brass filter, an exposure time of 32 ms, a voltage of 450 kV, and a current of 3000 µA, which were converted into 605 coronal slices with a voxel size of 500 µm. The specimens were visualized in 3-D using the Thermo Scientific Amira (6.4.0) software. The matrix was partially removed and sutures emphasized by adjusting thresholds manually.
The two shell specimens were photographed and illustrated using standard techniques in an attempt to capture their external morphology (Figs. 2, 3). Many sutures are not apparent externally, but clearly so in the CT images (Fig. 4). We therefore added to the illustrations sutures observed in the CT scans. These added sutures are highlighted in the illustrations by dashed lines to highlight their approximate nature (Figs. 2, 3).
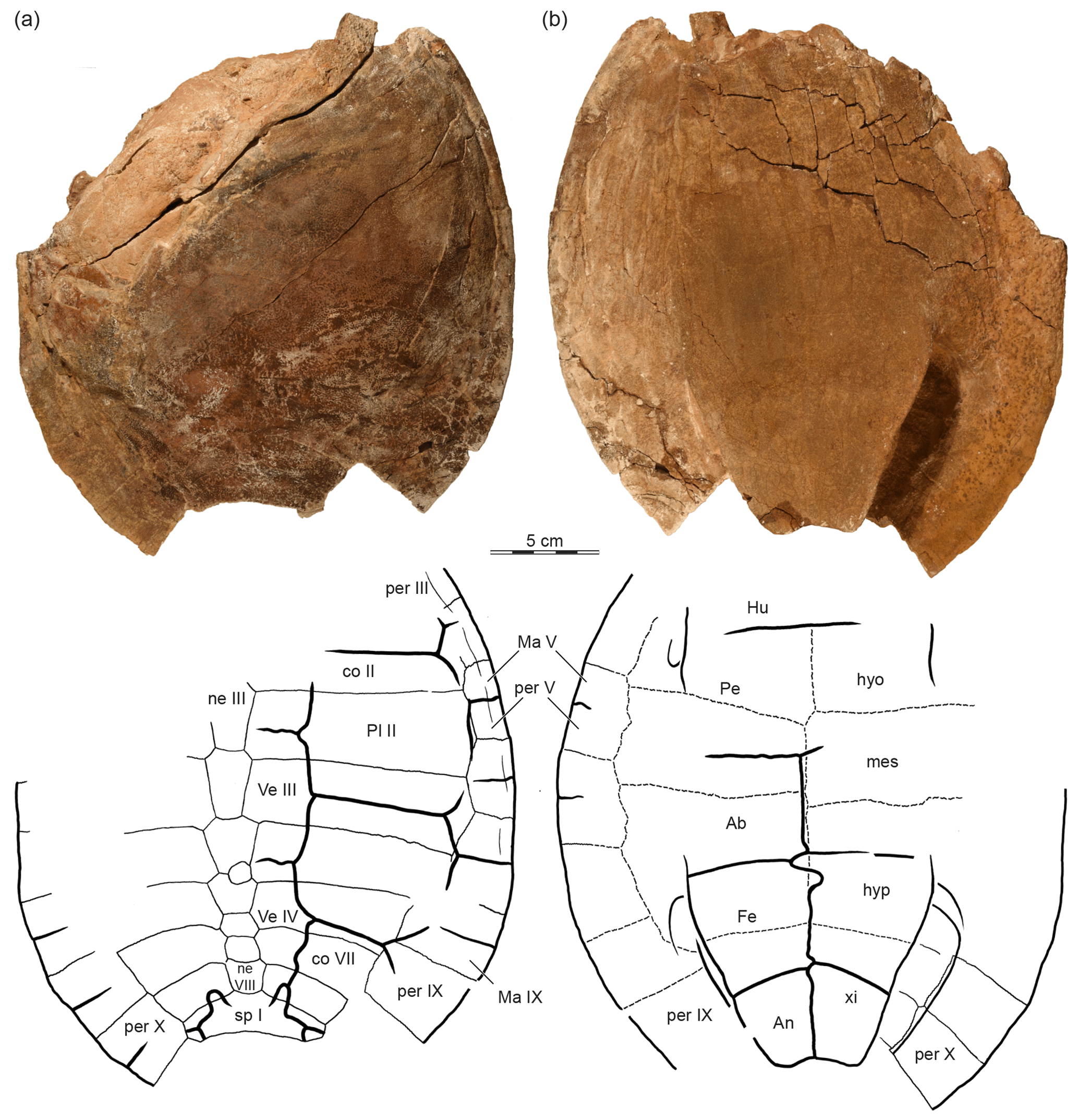
Figure 2OMNH 67133, a partial shell, holotype, Lakotemys australodakotensis gen. et. sp. nov., Berriasian–Barremian Lakota Formation, Fall River County, South Dakota, USA. Photographs and illustrations in (a) dorsal and (b) ventral view. Dashed lines connote sutures observed in 3-D-rendered CT scans (see Fig. 4). Abbreviations are as follows: Ab: abdominal scute; An: anal scute; co: costal; Fe: femoral scute; Hu: humeral scute; hyo: hyoplastron; hyp: hypoplastron; Ma: marginal scutes; mes: mesoplastron; ne: neural; Pe: pectoral scutes; per: peripheral; Pl: pleural scutes; sp: suprapygal; Ve: vertebral scute; xi: xiphiplastron.
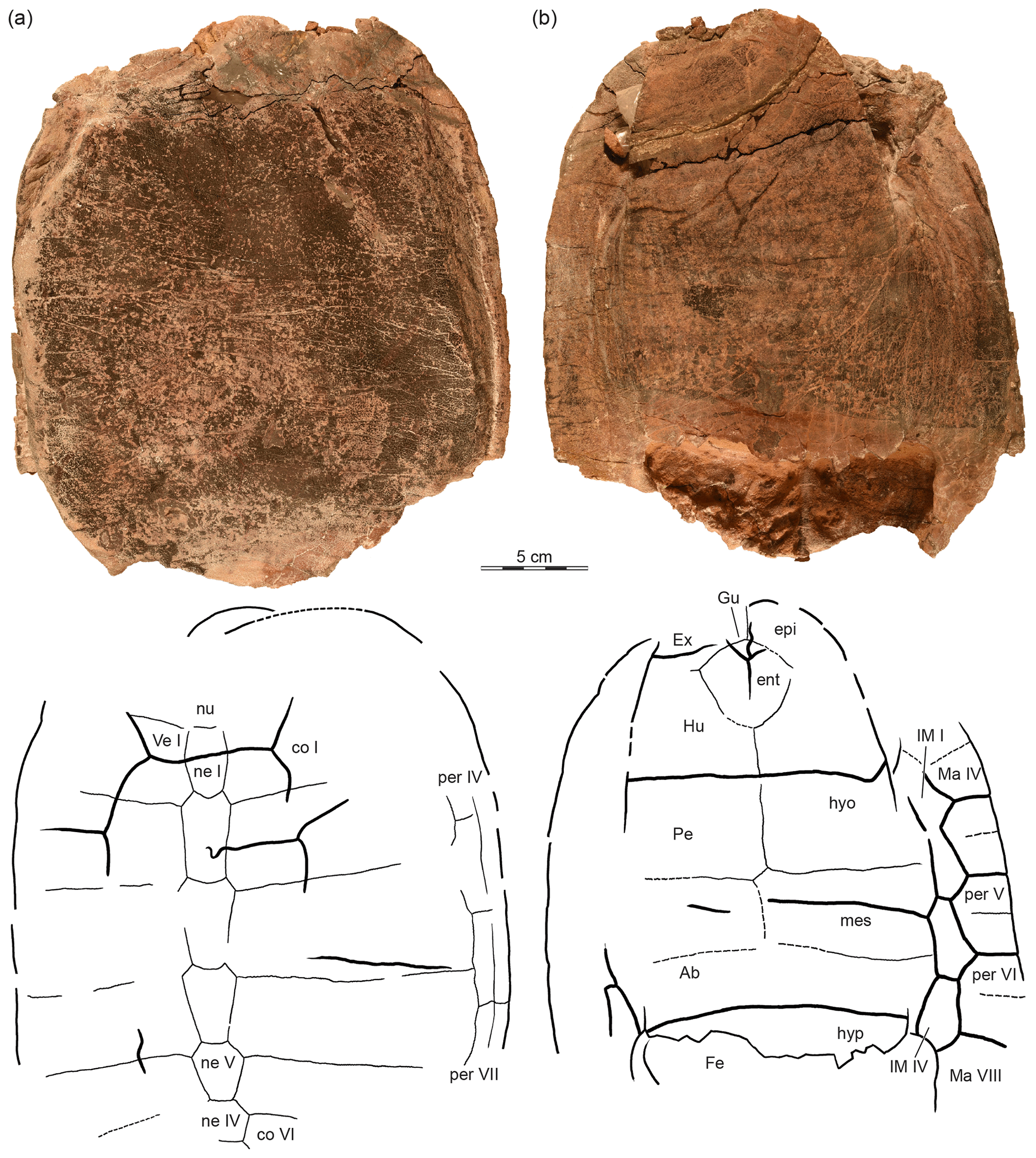
Figure 3OMNH 63615, a partial shell, Lakotemys australodakotensis gen. et. sp. nov., Berriasian–Barremian Lakota Formation, Fall River County, South Dakota, USA. Photographs and illustrations in (a) dorsal and (b) ventral view. Dashed lines connote sutures observed in 3-D-rendered CT scans (see Fig. 4). Abbreviations are as follows: Ab: abdominal scute; co: costal; ent: entoplastron; epi: epiplastron; Ex: extragular scute; Fe: femoral scute; Gu: gular scute; Hu: humeral scute; hyo: hyoplastron; hyp: hypoplastron; IM: inframarginal scute; Ma: marginal scutes; mes: mesoplastron; ne: neural; nu: nuchal scute; Pe: pectoral scutes; per: peripheral; Ve: vertebral scute.
The only available skull specimen is too poorly preserved to allow us to discern sutures either externally or in the CT scans. We therefore here only document the skull in its primary six views based on 3-D images obtained from the CT scans (Fig. 5).
-
Testudinata Batsch, 1788
-
Paracryptodira Gaffney, 1975
-
Baenidae Cope, 1873 (sensu Lyson and Joyce, 2011)
-
Lakotemys australodakotensis gen. et sp. nov.
Type specimen
OMNH 67133, a partial shell (Figs. 2, 4a).
Type locality and stratum
OMNH locality V1332, Dick Canyon, Fall River County, South Dakota (more detailed locality information is on file at OMNH); Unit 2, Chilson Member, Lakota Formation, Berriasian–Valanginian (see Geological settings above).
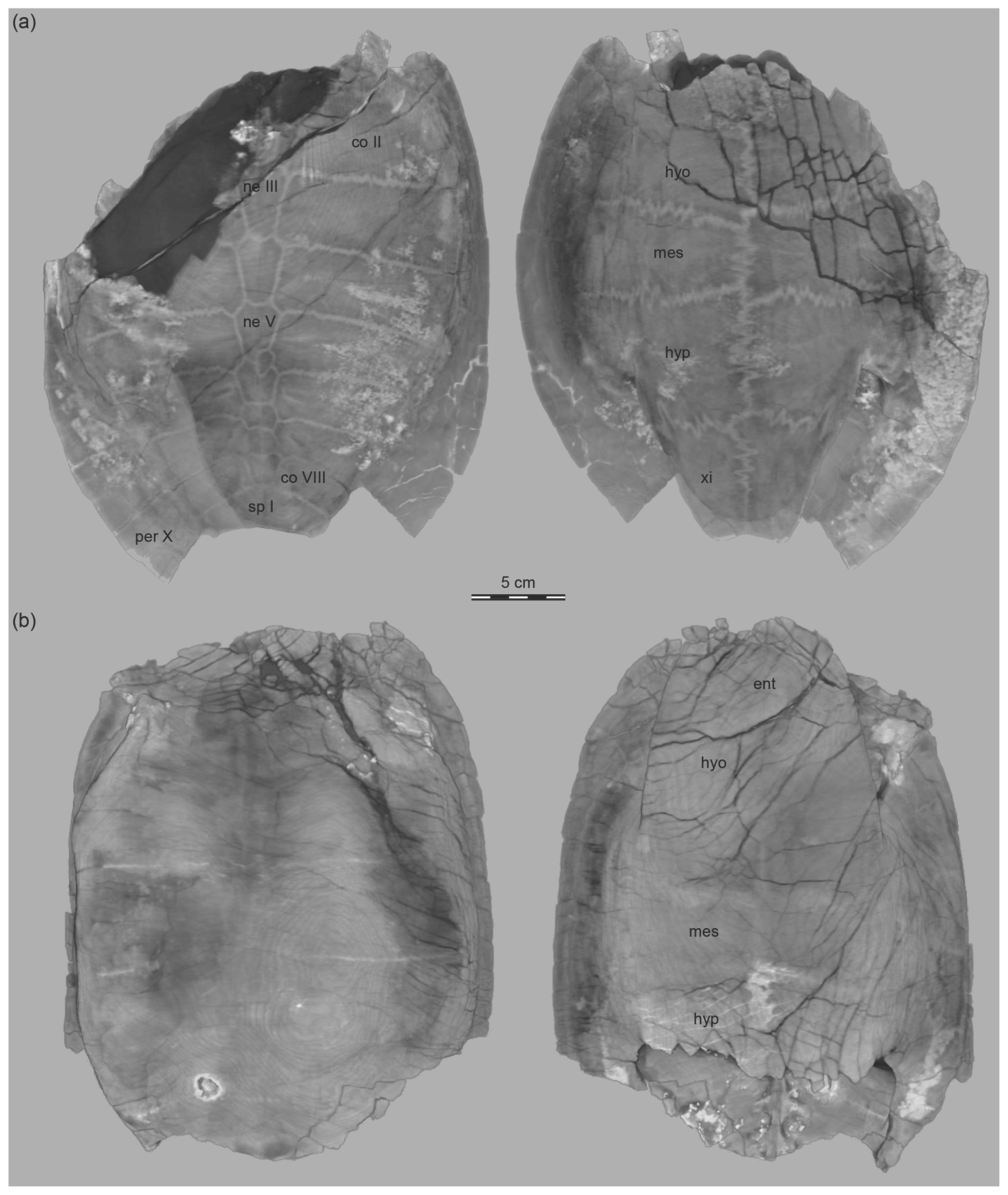
Figure 4The 3-D-rendered CT images of shells, Lakotemys australodakotensis gen. et. sp. nov., Berriasian–Barremian Lakota Formation, Fall River County, South Dakota, USA. Dorsal and ventral views of (a) OMNH 67133, the holotype, and (b) OMNH 63615. Abbreviations are as follows: co: costal; ent: entoplastron; hyo: hyoplastron; hyp: hypoplastron; mes: mesoplastron; ne: neural; per: peripheral; sp: suprapygal; xi: xiphiplastron.
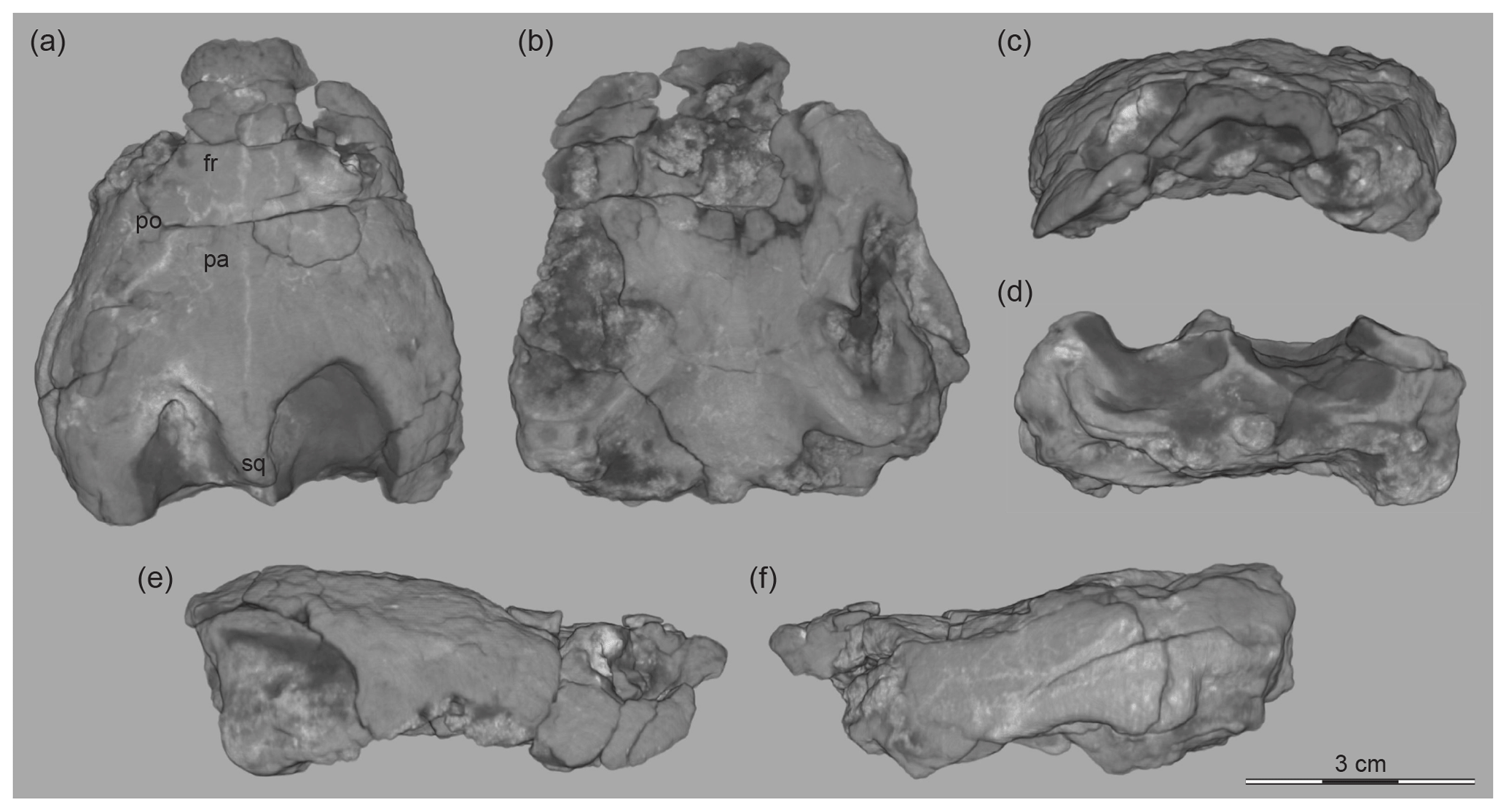
Figure 5The 3-D-rendered CT images of a skull, Lakotemys australodakotensis gen. et. sp. nov., Berriasian–Barremian Lakota Formation, Fall River County, South Dakota, USA. OMNH 66106 in (a) dorsal, (b) ventral, (c) anterior, (d) posterior, (e) right lateral, and (f) left lateral views. Abbreviations are as follows: fr: frontal; pa: parietal; po: postorbital.
Nomenclatural acts
This publication and its nomenclatural acts were registered at ZooBank on 12 February 2020, prior to publication. The LSID of the publication is urn:lsid:zoobank.org:pub:66C2A3D5-1E5C-42E8-8137-9B6253ACDA25, that of the new genus urn:lsid:zoobank.org:act:85E563EB-2F61-4560-8DB1-11F59261825E, and that of the new species urn:lsid:zoobank.org:act:00A9D083-7874-4022-AB3C-41B9FF71841E.
Diagnosis
Lakotemys australodakotensis gen. et sp. nov. can be diagnosed as a representative of Paracryptodira by the presence of a finely sculptured skull and shell and the likely location of the foramen posterius canalis carotici interni midline along the contact of the basisphenoid and pterygoid and as a representative of Baenidae by the absence of epiplastral processes and the development of well-developed axillary and inguinal buttresses. Among named Early Cretaceous baenids, Lakotemys australodakotensis can be differentiated from Protobaena wyomingensis and Trinitichelys hiatti by having an irregularly shaped vertebral V that does not lap onto neural VIII and that forms two anterolateral processes that partially hinder vertebral IV from contacting pleural IV and can be differentiated from Arundelemys dardeni and Trinitichelys hiatti by having a broader skull and triturating surfaces (see Discussion for additional differences).
Etymology
The compound word Lakotemys is derived from the combination of the word Lakota, in reference to the indigenous Lakota people of the greater Black Hills region, and the fossil-bearing Lakota Formation, and emys, ancient Greek for turtles. The species epithet australodakotensis is formed in allusion to the state of South Dakota through the combination of the word auster with the suffix -alis, Latin for south, and Dakota. The suffix -ensis is used to indicate a location.
Referred material
OMNH 66106, a poorly preserved skull collected from the type locality (Fig. 3); OMNH 63615, a poorly preserved shell collected from OMNH site V1382 (Fig. 4). All referred material originates from Unit 2 of the Chilson Member of the Lakota Formation.
Skull
The only available skull, OMNH 66106, is partially crushed and encrusted by a thick layer of metal oxides. The vast majority of sutures are obscured externally and in the CT scans. We therefore here only provide 3-D-rendered CT images of the skull in the primary six views and briefly highlight the most important features (Fig. 5).
Overall, the skull is characterized by being slightly longer than wide and by having intermediately developed lower and upper temporal emarginations, dorsoventrally oriented orbits, and a broad internarial bar. The size and orientation of the external nares are unclear, as is the size of the cavum tympani relative to the orbit. The overall shape nevertheless resembles the skull of most other baenids.
The frontal, postorbital, parietal, and squamosal are the only bones that can be at least partially recognized in the CT scans. The frontals and postorbitals broadly contribute to the dorsal margin of the orbit and therefore exclude the parietal from the orbit. The posterior tip of the short supraoccipital crest is not roofed by the parietal. As the remaining sutures are unclear, it is not possible to clarify on the dorsal skull surface the presence or size of nasals, the size of the dorsal lappets of the prefrontals, the contribution of the jugal to the orbit, or contact between the squamosal and parietal.
We are unable to discern any sutures on the ventral side of the skull. The anterior region is furthermore damaged, making it impossible to rigorously assess the morphology of the triturating surfaces. The posterior remnants of the maxilla nevertheless suggest that the triturating surfaces are expanded posteriorly. The external pterygoid processes are well developed and deeply protrude into the temporal fossa.
Despite the availability of CT scans, we are not able to trace the path of most cranial nerves and vessels. Faint traces of canals near the basisphenoid–pterygoid contact suggest that the internal carotid canal entered the skull halfway along the basisphenoid–pterygoid contact, but we cannot comment on the presence or absence of the palatal branch of the internal carotid or on the exact path of the facial nerve. The stapedial canals open into the upper temporal fossa on top of the otic capsule, but we are not able to clarify if the opisthotic contributes to its formation, both at the surface and within the skull.
Carapacial bones
Although both shells are incomplete, by reference to other paracryptodires we conclude with high confidence that they once consisted of a nuchal, eight neurals, two suprapygals, a pygal, eight pairs of costals, and 11 pairs of peripherals (Figs. 2, 3).
The nuchal is only partially preserved in OMNH 63615 and its preservation is poor (Fig. 3). The partially preserved anterior margin suggests that a distinct nuchal notch may have been present, while the partially preserved posterior margin highlights the contact of the nuchal with costal I. The likely lateral contact of the nuchal with peripheral I is not preserved.
OMNH 63615 preserves the anterior six neurals (Fig. 3), while OMNH 67133 preserves the posterior six neurals (Fig. 2). OMNH 67133 exhibits two minor abnormalities in the form of a doubled neural VII and the inclusion of an asymmetric ossification between neurals V and VI. The neurals form a continuous row of elements that fully separate the costals from one another. Neural I is rectangular and slightly shorter anteroposteriorly than neural II. The remaining neurals are hexagonal with short anterolateral sides. Although neurals II–VIII have a similar width, their anteroposterior lengths decrease gradually towards the posterior of the shell.
Suprapygal I is preserved in OMNH 67133 (Fig. 2). It is an octagonal element having a short anterior contact with neural VIII, an elongate anterolateral contact with costal VIII, a short lateral contact with peripheral X, a short posterolateral contact with peripheral XI, and a broad posterior contact with suprapygal II.
Suprapygal II is not preserved in either specimen, but the concave articular facet preserved along the posterior border of suprapygal I in OMNH 67133 (Fig. 2) is highly suggestive of its former presence. This element likely had a broad anterior contact with suprapygal I, a short posterolateral contact with peripheral XI, and a broad posterior contact with the pygal.
The pygal is not preserved in either specimen.
OMNH 67133 (Fig. 2) and OMNH 63615 (Fig. 3) combined document the presence of eight pairs of costals. From front to back, the costals gradually decrease in anteroposterior and mediolateral dimensions. While costal I medially contacts a single neural, all remaining elements are in contact with two neurals. In addition, costal I contacts the nuchal anteriorly, while costal VIII contacts suprapygal I posteriorly. The distal contacts of the costals are only partially preserved. Costal I likely contacted peripherals I–III. Costal II laterally contacts peripherals IV and V, costal III contacts peripherals V and VI, costal IV contacts peripherals VI and VII, costal V contacts peripherals VII and VIII, and costal VI contacts peripherals VIII and IX. Costal VII mostly contacts peripheral IX but also shows a minor contact with peripheral X. Finally, costal VIII only exhibits a lateral contact with peripheral X.
The specimens only preserve peripherals III–X, but the large gap at the back of OMNH 67133 (Fig. 2) suggests that 11 pairs were once present, instead of 10 pairs with an anomalously large pygal. Poor preservation obscures the morphology of the anterior peripherals, but peripherals I and II were likely flat. The bridge stretches from peripheral III to VIII. All bridge peripherals are laterally decorated by a well-developed, bead-like thickening. The posterior peripherals are once again flattened. Peripheral I likely contacted the nuchal medially. The likely or preserved medial contacts with the costals are described above. While peripheral X shares a short contact with suprapygal I, peripheral XI has a short contact with suprapygal I and likely has a broad anterior contact with suprapygal II and a medial contact with the pygal.
Carapacial scutes
OMNH 67133 and OMNH 63615 both preserve some traces of carapacial scutes, but as the observed pattern on the shell of OMNH 63615 is highly abnormal by showing pronounced asymmetry (Fig. 3), we restrict our description to the carapace of OMNH 67133 (Fig. 2).
Although only vertebrals II–V, pleurals I–IV, and marginals IV–X are preserved, the shell was likely covered by five vertebrals, four pairs of pleurals, and 12 marginals. The former presence of cervicals cannot be ascertained as the surface of the nuchal is poorly preserved.
Vertebral II is only partially preserved but appears to have similar dimensions to that of vertebral III (Fig. 2). Vertebral III is six-sided, but the parallel lateral margins give it an overall square outline. Vertebral III contacts vertebral II anteriorly along a sulcus that crosses neural III, pleurals II and III laterally, and vertebral IV posteriorly along a sulcus that crosses neural V. Vertebral IV is approximately as wide as vertebral III, but is slightly longer and has a more hexagonal outline. In addition to contacting vertebral III anteriorly, it contacts pleurals III and IV laterally and forms a highly sinuous posterior sulcus with vertebral V that posteriorly circumnavigates neural VIII to cross the suprapygal. Only the anterior half of vertebral V is preserved. This portion of this scute documents a highly unusual morphology consisting of two fingerlike projections that intrude between vertebral IV and pleural IV. The highly symmetric arrangement suggests that this is not an irregularity. The sulcus with pleural IV is otherwise located on suprapygal I. As the posterior elements are not preserved, it is not clear if vertebral V contributed to the posterior margin of the carapace, the condition seen in baenodds. However, the plesiomorphic, expanded nature of the most posteriorly preserved peripherals makes this unlikely.
The pleurals are slightly wider than the vertebrals (Fig. 2). The interpleural sulci are straight and located on costals II, IV, and VI. Pleural I at least contacted marginal IV and V laterally, pleural II contacted marginals V–VII, pleural III contacted marginals VII–IX, and pleural IV contacted marginals IX–XI. The contacts of marginals I–III are not preserved. The marginals alternatingly have rectangular to pentagonal outlines. The tips of the pentagons partially subdivide the pleurals and rise to the level of the costals.
Plastral bones
The anterior plastral lobe is better preserved in OMNH 63615 (Fig. 3), and the posterior lobe is better preserved in OMNH 67133 (Fig. 2). The two specimens therefore jointly provide a nearly complete picture of this part of the shell.
The plastron consists of an entoplastron and pairs of epi-, hyo-, meso-, hypo-, and xiphiplastra. The arrangement and contacts of these bones resemble those of other turtles. The epiplastra are notable by having a broad midline contact. The anterior portion of the large entoplastron is smaller than the posterior portion. The anterior plastral lobe is broad, elongate, and rounded and mostly formed by the enlarged hyoplastra. It likely protruded anteriorly beyond the anterior margin of the carapace. The hyo-, meso-, and hypoplastral jointly form the bridge that articulates with peripherals III–VIII. The exact nature of the buttress is obscured by matrix but can nevertheless be ascertained to be more expansive than in Glyptops ornatus, by articulating with the costals. The mesoplastra are large elements that form an irregular, offset contact with one another and the neighboring elements along the midline. They laterally contact peripherals V and VI. The posterior plastral lobe is shorter and narrower than the anterior lobe, is constricted towards the posterior, and only partially covers the carapace. It is jointly formed by the hypoplastra and xiphiplastra. An anal notch is present but shallow.
Plastral scutes
The plastron is covered by pairs of gulars, extragulars, humerals, pectorals, abdominals, femorals, and anals and four pairs of inframarginals. The gulars and extragulars are only partially preserved, but it is possible to ascertain that the gulars were paired, triangular elements that had a broad midline contact and lapped onto the entoplastron and that the extragulars did not have a midline contact or lap onto the entoplastron. The extragular–humeral sulcus is located on the epiplastron, the humeral–pectoral sulcus on the hyoplastron just anterior to the axillary notch, the pectoral/abdominal sulcus on the mesoplastra, the abdominal–femoral sulcus on the hypoplastra just posterior to the inguinal notch, and the femoral–anal sulcus on the xiphiplastra far from its contact with the hypoplastron. The pectoral and abdominal scutes form the deepest portions of the axillary and inguinal notches, respectively. Inframarginal I medially contacted the pectoral and laterally contacted at least marginals IV and V.
We inserted Lakotemys australodakotensis gen. et sp. nov. into the matrix of Lyson et al. (2019), which in turn is based on the phylogenetic analysis of Lyson and Joyce (2009a, b, 2010, 2011), Lyson et al. (2011), and Lyson et al. (2016). To investigate the phylogenetic relationships of Early Cretaceous paracryptodires, we furthermore included Protobaena wyomingensis from the Aptian–Albian of Wyoming to the matrix, based on the original description of Gilmore (1919). The final character–taxon matrix consisting of 36 taxa scored for 69 characters is provided in the Supplement.
The matrix was subjected to a parsimony analysis using TNT (Goloboff et al., 2008). Twelve characters form morphoclines and were run ordered (i.e., characters 4, 10, 16, 21, 22, 23, 25, 29, 34, 40, 44, 45). Following the recommendation of Goloboff et al. (2018), light implied weighting was implemented using a K factor of 12. The matrix was subjected to 1000 replicates of random addition sequences followed by a second round of tree bisection and reconnection. The primary analysis using all taxa and characters resulted in five most parsimonious trees with a best score of 6.80893. The prune tree function of TNT identified Arundelemys dardeni as the only rogue taxon. We therefore ran a secondary analysis omitting this fossil turtle, resulting in a single most parsimonious tree with a best score of 6.76481. The strict consensus tree resulting from the omission of Arundelemys dardeni post-analysis is identical to the single tree retrieved from the omission of Arundelemys dardeni pre-analysis (Fig. 6).
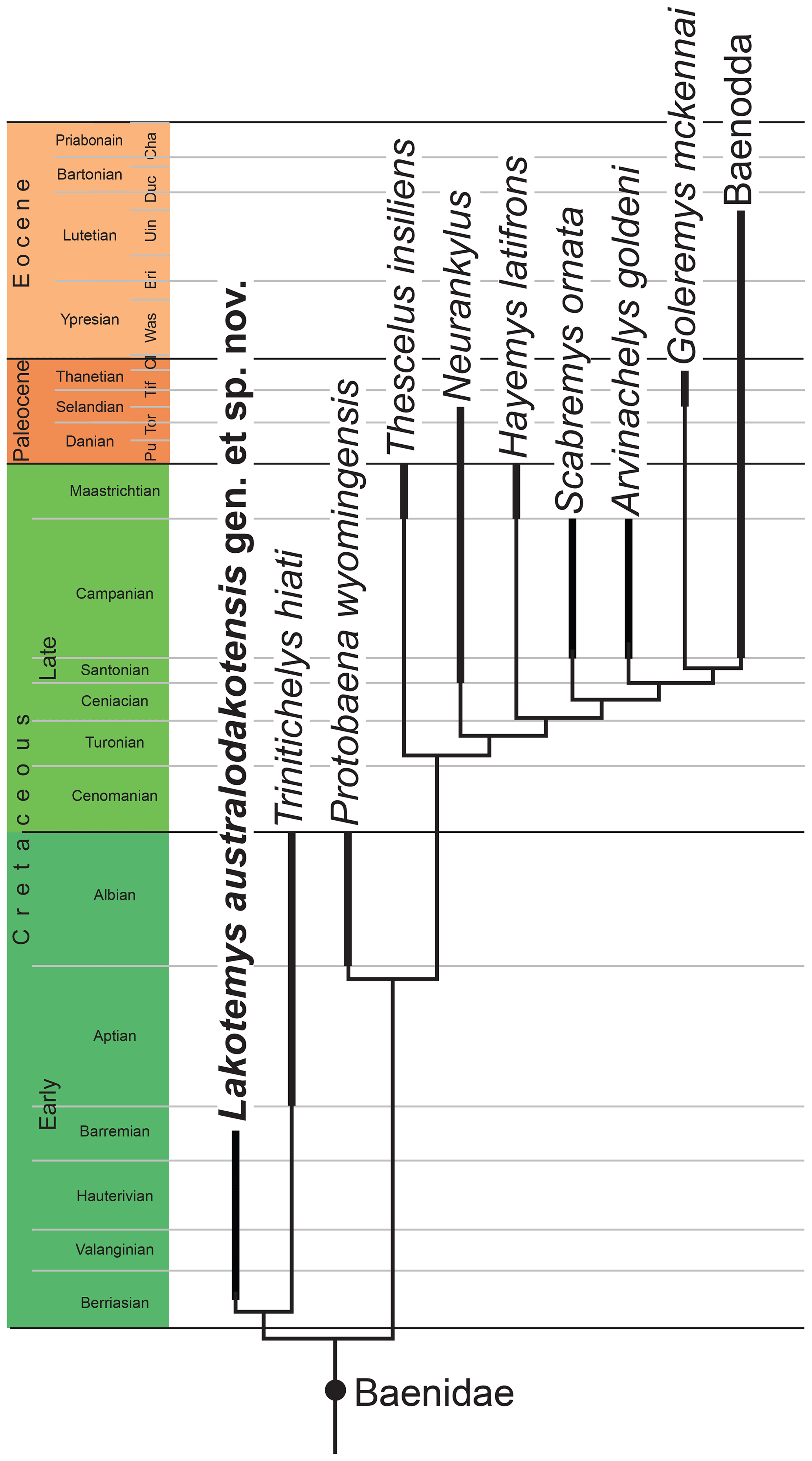
Figure 6Time-calibrated strict consensus topology obtained from both phylogenetic analyses. Out-groups are removed and all derived baenids united into the clade Baenodda.
Alpha taxonomy
We here refer three specimens to Lakotemys australodakotensis gen. et sp. nov. The holotype, a partial shell, and the only known skull were collected from the same quarry and may reasonably originate from the same specimen, but this cannot be ascertained with confidence. The second referred specimen, a shell, was collected from nearby outcrops of the same unit within the Lakota Formation. The three specimens therefore at least represent a single species, at most three species. The hypothesis that all three fossils represent a single species is supported by common provenance (the same rock layer and/or quarry), shared characteristics (overall similarity in shell morphology), and the realization that other Early Cretaceous formations are not known to be particularly diverse. The two major conclusions drawn herein, in particular the naming of a new species and its identification as a basal baenid, however, should remain unaffected if future finds indicate that the three specimens represent separate species. Indeed, as the skull and referred shell are not particularly informative, both conclusions can be justified on the basis of OMNH 67133 (the holotype) alone, as this specimen exhibits the majority of informative character information.
We summarize in the following section differences with all other currently recognized Late Jurassic to Early Cretaceous paracryptodires worldwide (Joyce and Lyson, 2015; Joyce and Anquetin, 2019).
The Late Jurassic (Kimmeridgian) Selenemys lusitanica Pérez-García and Ortega, 2011 from Portugal substantially differs from Lakotemys australodakotensis gen. et sp. nov. by having a more coarsely textured shell; a short, midline contact of peripheral 1 anterior to the nuchal; a neural reversal; a suprapygal I that is much smaller than suprapygal II; broader and shorter plastral lobes; a much larger entoplastron; broader, regularly shaped and positioned vertebrals; and a highly sinuous midline plastral sulcus.
The Late Jurassic (Tithonian) Glyptops ornatus Marsh, 1890 from North America, as described by Hay (1908) and Gaffney (1979) under the name Glyptops plicatulus, resembles Lakotemys australodakotensis in possessing a finely textured skull and shell, and in the overall configuration of the shell, but differs by being smaller, having a much more elongate skull with delicate triturating surfaces; a smaller suprapygal I that does not contact the peripherals; a shorter, but broader anterior plastral lobe; and regularly positioned, hexagonal vertebral scutes.
The Late Jurassic (Tithonian) Dinochelys whitei Gaffney, 1979 from North America differs substantially from Lakotemys australodakotensis by lacking a textured shell; by having a neural reversal; by lacking a contact of suprapygal I with the peripherals; by having much broader, regularly shaped and positioned vertebrals and shorter plastral lobes; by lacking an overlap of the gulars onto the entoplastron; and by having an overlap of the anal onto the hypoplastron.
The Late Jurassic (Tithonian) Riodevemys inumbragigas Pérez-García et al., 2015b from Spain resembles Lakotemys australodakotensis in possessing a finely textured skull and shell, but differs by having a smaller suprapygal I than suprapygal II, broader and shorter plastral lobes, and a much larger entoplastron; by lacking a midline contact of the mesoplastra; and by having broader, regularly shaped and positioned vertebrals.
The Early Cretaceous (Berriasian) Pleurosternon bullockii (Owen, 1842) from the UK, as described by Evans and Kemp (1975) and Milner (2004), resembles Lakotemys australodakotensis in possessing a finely textured skull and shell, but differs by having a much more elongate skull with narrow triturating surfaces, a suprapygal I that is much smaller than suprapygal II, wider, but regularly positioned vertebrals, a broader anterior plastral lobe, a much larger entoplastron, and a distinct anal notch.
The Early Cretaceous (Berriasian) Pleurosternon typocardium (Seeley, 1869) from western Europe, as described by Evans and Kemp (1976) and Pérez-García (2014), resembles Lakotemys australodakotensis in the overall gestalt and configuration of the skull and shell, and by having a lateral contact of suprapygal I with the peripheral series, but differs by having a neural reversal, regularly shaped and positioned vertebrals, a shorter anterior plastral lobe, a much larger entoplastron, and, likely, a deep nuchal notch.
The Early Cretaceous (Albian) Toremys cassiopeia Pérez-García et al., 2015a from Spain resembles Lakotemys australodakotensis by having finely textured shell sculpturing, but differs by being much smaller and having a shorter anterior plastral lobe, a large entoplastron, and a deeper anal notch.
The Early Cretaceous (Aptian–Albian) Protobaena wyomingensis (Gilmore, 1919) from Wyoming resembles Lakotemys australodakotensis in having a finely textured shell surface, but differs by having regularly shaped and placed vertebral scutes, a pygal notch that reaches peripheral X, and a narrower posterior plastral lobe.
The Early Cretaceous (Aptian–Albian) Trinitichelys hiatti Gaffney, 1972 from Texas resembles Lakotemys australodakotensis by having a finely textured shell surface, the proportions of the plastral lobes, and the overall arrangement of bones and scutes, but differs by having a more elongate skull and less distance between the triturating surfaces, regularly placed vertebral scutes with sinuous margins, and a greater overlap of the gulars onto the entoplastron.
The Late Jurassic (Tithonian) Uluops uluops Carpenter and Bakker, 1990 and the Aptian–Albian Arundelemys dardeni Lipka et al., 2006 are only known from cranial material. The poor preservational state of the only available skull of Lakotemys australodakotensis therefore makes it difficult to highlight meaningful differences. We nevertheless note that the skulls of Uluops uluops and Arundelemys dardeni are more elongate and have much narrower triturating surfaces. In addition, the ventral side of the basisphenoid of Uluops uluops is decorated with processes, while the pterygoid of Arundelemys dardeni exhibits deep pterygoid fossae.
In conclusion, the new turtle material documents a unique morphotype and we therefore feel justified designating it as representing a new species, Lakotemys australodakotensis gen. et sp. nov.
Phylogenetic relationships
Our phylogenetic analysis hypothesizes that the early Early Cretaceous Lakotemys australodakotensis gen. et sp. nov. is the immediate sister of the late Early Cretaceous Trinitichelys hiatti at the base of Baenidae. The late Early Cretaceous Protobaena wyomingensis is the next more highly nested representative. This conclusion overall suggests that Early Cretaceous baenids form a lineage that gradually acquired characters more typical of Late Cretaceous and Paleogene forms. The placement of Lakotemys australodakotensis and Trinitichelys hiatti at the base of Baenidae is suggested by the presence of an extended midline contact of the pterygoids (not preserved in Lakotemys australodakotensis), absence of epiplastral processes, and the development of well-developed axillary and inguinal buttresses. The sister-group relationship of Lakotemys australodakotensis and Trinitichelys hiatti is not actually supported by a character, but rather appears to be a result of the use of implied weighting. More derived characters that unite Protobaena wyomingensis with all Late Cretaceous to Paleogene baenids include the fusion of the shell, but we also note that Protobaena wyomingensis has an incipient pygal notch, which appears intermediate to the deep pygal notch of baenodds.
Late Cretaceous baenids have already historically been thought to be closely related with Late Jurassic pleurosternids (e.g., Hay, 1908). This sister-group relationship was supported by more recent phylogenies as well (e.g., Joyce, 2007; Lyson and Joyce, 2011), but it also suggests an undocumented baenid ghost lineage that crosses the early Early Cretaceous into the Late Jurassic. The herein described material from the early Early Cretaceous (late Berriasian–early Barremian) Lakota Formation of South Dakota helps to partially fill this gap. We nevertheless note that the earliest-known pleurosternids are Kimmeridgian in age, which suggests that baenids must have existed during the deposition of the Late Jurassic Morrison Formation as well. The origin of the baenid lineage is therefore to be expected among Morrison fossils previously described as “glyptopsids”.
The CT slice data produced for this study are permanently archived at the University of Texas High-Resolution X-ray Computed Tomography Facility and available upon request. The character–taxon matrix is available in the Supplement.
The supplement related to this article is available online at: https://doi.org/10.5194/fr-23-1-2020-supplement.
The material described herein was collected by RLC. WGJ and YR photographed, illustrated, and described the material, performed the phylogenetic analysis, and wrote the primary draft of the manuscript. RLC wrote the geological background. All authors read and approved the final draft.
The authors declare that they have no conflict of interest.
We thank Susan Watts and the late Dayton Hyde for allowing RLC access to the Black Hills Wild Horse Sanctuary. It is also a pleasure to acknowledge, with thanks, Jim Angell and Karen Hossack, for their hospitality and help to OMNH field crews through the years. We also thank Joshua Lively, Johannes Müller, and Nicole Klein for providing insightful comments that helped improve the quality of the manuscript.
This research has been supported by the Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (grant no. 200021_178780/1).
This paper was edited by Johannes Müller and reviewed by Joshua Lively and Nicole Klein.
Batsch, A. J. G. C.: Versuch einer Anleitung, zur Kenntniß und Geschichte der Thiere und Mineralien. Jena: Akademische Buchhandlung, 528 pp., 1788.
Bell, H. and Post, E. V.: Geology of the Flint Hill Quadrangle, Fall River County, South Dakota, U. S. Geol. Surv. Bull., 1063-M, 505–586, 1971.
Bolyard, D. W. and McGregor, A. A.: Stratigraphy and petroleum potential of Lower Cretaceous Inyan Kara Group in northeastern Wyoming, southeastern Montana, and western South Dakota, Bull. Am. Ass. Petrol. Geol., 50, 2221–2244, 1966.
Bonde, J. W., Varricchio, D. J., Jackson, F. D., Loope, D. B., and Shirk, A. M.: Dinosaurs and dunes! Sedimentology and paleontology of the Mesozoic in the Valley of Fire State Park, Geol. Soc. Am. Field Guide, 11, 249–262, 2008.
Brinkman, D. B.: A review of nonmarine turtles from the Late Cretaceous of Alberta, Can. J. Earth Sci., 40, 557–571, 2003.
Brinkman, D. B.: Turtles: diversity, paleoecology and distribution, in: Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed, edited by: Currie, P. J. and Koppelhus, E. B., Indiana University Press, Bloomington, Indiana, 202–202, 2005.
Carpenter, K. and Bakker, R. T.: A new latest Jurassic vertebrate fauna, from the highest levels of the Morrison Formation at Como Bluff, Wyoming, with comments on Morrison biochronology, Part II. A new baenid turtle, Hunteria, 2, 1–6, 1990.
Cifelli, R. L., Gardner, J. D., Nydam R. L., and Brinkman, D. L.: Additions to the vertebrate fauna of the Antlers Formation (Lower Cretaceous), southeastern Oklahoma, Okla. Geol. Notes, 57, 124–131, 1997.
Cifelli, R. L., Nydam, R. L., Gardner, J. D., Weil, A., Eaton, J. G., Kirkland, J. I., and Madsen, S. K.: Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County, Utah: the Mussentuchit Local Fauna, in: Vertebrate Paleontology in Utah, edited by: Gillette, D. D., Utah Geological Survey, Salt Lake City, 219–242, 1999.
Cifelli, R. L., Davis, B. M., and Sames, B.: Earliest Cretaceous mammals from the western United States, Act. Palaeontol. Pol., 59, 31–52, 2014.
Cope, E. D.: Descriptions of some new Vertebrata from the Bridger Group of the Eocene, Proc. Am. Phil. Soc., 12, 460–465, 1873.
Dahlstrom, D. J. and Fox, J. E.: Fluvial architecture of the Lower Cretaceous Lakota Formation, southwestern flank of the Black Hills Uplift, South Dakota, B. U. S. Geol. Surv., 1917, S1–S20, 1995.
Dandavati, K. S.: Continental and near-shore depositional environments of the Inyan Kara Group, southeastern Black Hills, South Dakota, in: Geology of the Black Hills, South Dakota and Wyoming: Fieldtrip Guidebook, Geological Society of America, 19–31, 1985.
Darton, N. H.: Preliminary description of the geology and water resources of the southern half of the Black Hills and adjoining regions in South Dakota and Wyoming, An. Rep. U. S. Geol. Surv., 21, 489–599, 1901.
DeCelles, P. G.: Late Jurassic to Eocene evolution of the Cordilleran thrust belt and foreland basin system, Western USA, Am. J. Sci., 304, 105–168, 2004.
D'Emic, M. D., Foreman, B. Z., Jud, N. A., Britt, B. B., Schmitz, M., and Crowley, J. L.: Chronostratigraphic revision of the Cloverly Formation (Lower Cretaceous, Western Interior, USA), Bull. Peabody Mus. Nat. Hist., 60, 3–40, 2019.
Evans, J. and Kemp, T. S.: The cranial morphology of a new Lower Cretaceous turtle from Southern England, Palaeontol., 18, 25–40, 1975.
Evans, J. and Kemp, T. S.: A new turtle skull from the Purbeckian of England and a note on the early dichotomies of cryptodire turtles, Palaeontol., 19, 317–324, 1976.
Gaffney, E. S.: The systematics of the North American family Baenidae (Reptilia, Cryptodira), Bull. Am. Mus. Nat. Hist., 147, 241–320, 1972.
Gaffney, E. S.: A phylogeny and classification of higher categories of turtles, Bull. Am. Mus. Nat. Hist., 155, 387–436, 1975.
Gaffney, E. S.: The Jurassic turtles of North America, Bull. Am. Mus. Nat. Hist., 162, 91–136, 1979.
Gilmore, C. W.: New fossil turtles, with notes on two described species, Proc. U. S. Nat. Mus., 56, 113–132, 1919.
Goloboff, P. A., Farris, J., and Nixon, K.: TNT: a free program for phylogenetic analysis, Cladistics, 24, 774–786, 2008.
Goloboff, P. A., Torres, A., and Salvador Arias, J.: Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology, Cladistics, 34, 407–437, 2018.
Gott, G. B., Wolcott, D. E., and Bowles, C. G.: Stratigraphy of the Inyan Kara Group and localization of uranium deposits, southern Black Hills, South Dakota and Wyoming, Geol. Surv. Prof. Pap., 763, 1–57, 1974.
Hay, O. P.: The Fossil Turtles of North America, Carn. Inst. Wash. Pub., 75, 1–568, 1908.
Joyce, W. G.: Phylogenetic relationships of Mesozoic turtles, Bull. Peabody Mus. Nat. Hist., 48, 3–102, 2007.
Joyce, W. G. and Anquetin, J.: A review of the fossil record of non-baenid turtles of the clade Paracryptodira, Bull. Peabody Mus. Nat. Hist., 60, 129–155, 2019.
Joyce, W. G. and Lyson, T. R.: A review of the fossil record of turtles of the clade Baenidae, Bull. Peabody Mus. Nat. Hist., 56, 147–183, 2015.
Lipka, T. R., Therrien, F., Weishampel, D. B., Jamniczky, H. A., Joyce, W. G., Colbert, M. W., and Brinkman, D. B.: A new turtle from the Arundel Clay Facies (Potomac Formation, Early Cretaceous) of Maryland, USA, J. Vert. Paleontol., 26, 300–307, 2006.
Lyson, T. R. and Joyce, W. G.: A new species of Palatobaena (Testudines: Baenidae) and a maximum parsimony and Bayesian phylogenetic analysis of the Baenidae, J. Paleontol., 83, 457–470, 2009a.
Lyson, T. R. and Joyce, W. G.: A revision of Plesiobaena (Testudines: Baenidae) and an assessment of baenid ecology across the K/T boundary, J. Paleontol., 83, 833–853, 2009b.
Lyson, T. R. and Joyce, W. G.: A new baenid turtle from the Upper Cretaceous (Maastrichtian) Hell Creek Formation of North Dakota and a preliminary taxonomic review of Cretaceous Baenidae, J. Vert. Paleontol., 30, 394–402, 2010.
Lyson, T. R. and Joyce, W. G.: Cranial anatomy and phylogenetic placement of the enigmatic turtle Compsemys victa Leidy, 1856, J. Paleontol., 85, 789–801, 2011.
Lyson, T. R., Joyce, W. G., Lucas, S. G., and Sullivan, R. M.: A new baenid turtle from the early Paleocene (Torrejonian) of New Mexico and a species level phylogenetic analysis of Baenidae, J. Paleontol., 90, 305–316, 2016.
Lyson, T. R., Sayler, J. L., and Joyce, W. G.: Anew baenid turtle, Saxochelys gilberti, gen. et sp. nov., from the uppermost Cretaceous (Maastrichtian) Hell Creek Formation: sexual dimorphism and spatial niche partitioning within the most speciose group of Late Cretaceous turtles, J. Vert. Paleontol., 39, e1662428, https://doi.org/10.1080/02724634.2019.1662428, 2019.
Marsh, O. C.: Notice of some extinct Testudinata, Am. J. Sci., 40, 177–179, 1890.
Milner, A. R.: The turtles of the Purbeck Limestone Group of Dorset, southern England, Palaeontol., 47, 1441–1467, 2004.
Oreska, M. P. J., Carrano, M. T., and Dzikiewicz, K. M.: Vertebrate paleontology of the Cloverly Formation (Lower Cretaceous), I: faunal composition, biogeographic relationships, and sampling, J. Vert. Paleontol., 33, 264–292, 2013.
Owen, R.: Report on British fossil reptiles, Part II, Rep. Brit. Ass. Adv. Sci., 11, 60–204, 1842.
Pérez-García, A.: Revision of the poorly known Dorsetochelys typocardium, a relatively abundant pleurosternid turtle (Paracryptodira) in the Early Cretaceous of Europe, Cret. Res., 49, 152–162, 2014.
Pérez-García, A. and Ortega, F.: Selenemys lusitanica, gen. et sp. nov., a new pleurosternid turtle (Testudines: Paracryptodira) from the Upper Jurassic of Portugal, J. Vert. Paleontol., 31, 60–69, 2011.
Pérez-García, A., Espílez, E., Mampel, L., and Alcalá, L.: A new European Albian turtle that extends the known stratigraphic range of the Pleurosternidae (Paracryptodira), Cret. Res., 55, 74–83, 2015a.
Pérez-García, A., Royo-Torres, R., and Cobos, A.: A new European Late Jurassic pleurosternid (Testudines, Paracryptodira) and a new hypothesis of paracryptodiran phylogeny, J. Syst. Palaeontol., 13, 351–369, 2015b.
Post, E. V.: Geology of the Cascade Springs Quadrangle, Fall River County, South Dakota, U. S. Geol. Surv. Bull., 1063-L, 443–504, 1967.
Sames, B.: Taxonomic studies in Early Cretaceous nonmarine Ostracoda of North America, Micropaleontol, 57, 289–465, 2011.
Sames, B., Cifelli, R. L., and Schudack, M. E.: The nonmarine Lower Cretaceous of the North American Western Interior foreland basin: new biostratigraphic results from ostracod correlations and early mammals, and their implications for paleontology and geology of the basin – an overview, Earth Sci. Rev., 101, 207–224, 2010.
Seeley, H. G.: Index to the Fossil Remains of Aves, Ornithosauria, and Reptilia, from the Secondary System of Strata Arranged in the Woodwardian Museum of the University of Cambridge, Deighton, Bell, Cambridge, UK, 143 pp., 1869.
Sohn, I. G.: Nonmarine ostracodes in the Lakota Formation (Lower Cretaceous) from South Dakota and Wyoming, Geol. Surv. Prof. Pap., 1069, 1–22, 1979.
Szigeti, G. J. and Fox, J. E.: Unkpapa Sandstone (Jurassic), Black Hills, South Dakota: an eolian facies of the Morrison Formation, in: Recent and Ancient Nonmarine Depositional Environments: Models for Exploration, edited by: Ethridge, F. G. and and Flores, R. M., SEPM Society for Sedimentary Geology, Tulsa, Oklahoma, 331–349, 1981.
Waagé, K. M.: Stratigraphy of the Inyan Kara Group in the Black Hills, U. S. Geol. Surv. Bull., 1081-B, 11–90, 1959.
Walker, J. D., Geissman, J., Bowring, S. A., and Babcock, L. E.: Geologic Time Scale v. 5.0, Geological Society of America, 2018.
Ward, L. F.: The Cretaceous rim of the Black Hills, J. Geol., 2, 250–266, 1894.
Way, J. N., O'Malley, P. J., Suttner, L. J., and Furer, L. C.: Tectonic controls on alluvial systems in a distal foreland basin: the Lakota and Cloverly formations (Early Cretaceous) in Wyoming, Montana and South Dakota, Sp. Pub. Soc. Sed. Geol., 59, 133–147, 1998.
Zaleha, M. J.: Sevier orogenesis and nonmarine basin filling: implications of new stratigraphic correlations of Lower Cretaceous strata throughout Wyoming, USA, Bull. Geol. Soc. Am., 118, 886–896, 2006.